T. H. Huxley napsal, že dívat se na jiné lidoopy znamená vidět „rozmazané kopie sebe sama“ (1863 str. 73). Blízkou fyzickou příbuznost mezi opicemi neboli hominoidy rozpoznal již o století dříve Carolus Linnaeus (1759). Žijící hominoidy spojují znaky související s obvyklou ortografií a podbřišním chováním: široký trup s široce rozkročenými ramenními klouby, tuhá záda, dlouhé přední končetiny, pohyblivé klouby končetin, silná úchopová schopnost a absence ocasu (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Lidé některé z těchto znaků, například relativně dlouhé paže, postrádají v důsledku sekundární adaptace na bipedalismus. Také lebeční atributy jsou hominoidům společné, včetně širokého předního patra a relativně velkého mozku (Jerison, 1973; Rae, 1997). Hominoidi mají také prodlouženou délku života s opožděným dospíváním (Kelley, 1997) a zaujímají podobné ekologické niky – všechny druhy kromě člověka obývají tropické lesy a jsou významně závislé na zralých plodech a listech jako potravě (Fleagle, 2013).
Hylobatidi (giboni a siamangové) jsou menší než ostatní žijící lidoopi a liší se i v dalších ohledech, což odráží jejich dlouhé období evoluční izolace od hominidů (lidoopů a lidí), kteří mají společného nedávného předka (obr. 1). Hominidé (kromě člověka) mají přední patro rozšířené více než hylobatidé (Rae, 1997), mají ještě kratší a tužší spodní část zad a postrádají anatomické specializace pro ricochetální brachializaci (Young, 2003).
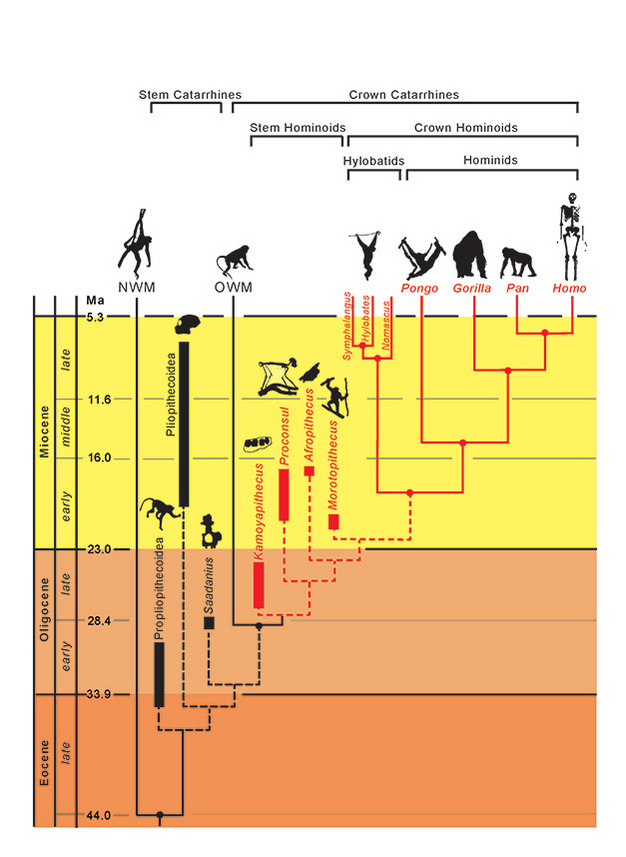
Fylogenetické vztahy u taxonů jsou znázorněny plnými (existující) a čárkovanými (vymřelé) čarami. Geologický čas je znázorněn na svislé ose. Doba větvení taxonů je pouze přibližná a odvozená z kombinace molekulárních a fosilních dat (Steiper et al., 2004; MacLatchy et al., 2006; Steiper a Young, 2008 a 2009; Chan et al., 2010; Stevens et al., 2013). Fylogenetické zařazení fosilních taxonů je hypotetické, jak je uvedeno v textu. Pozice Afropithecus a Morotopithecus může být obrácená, nebo mohou být oba taxony sesterské. NMW se vztahuje k opicím Nového světa a OWM se vztahuje k opicím Starého světa, zatímco Propliopithecoidea, Pliopithecoidea a Saadanius se vztahují ke kmenovým katarrhinům z Afroarabie postrádajícím jakékoliv synapomorfie korunových katarrhinů.
Vzhledem k těmto adaptivním podobnostem mezi žijícími hominoidy je zajímavé zkoumat, které (pokud vůbec nějaké) charakterizovaly počáteční divergenci hominoidů od ostatních katarrhinních primátů, zejména cercopithecoidů (opic Starého světa). Kombinované morfologické a genetické důkazy kladou divergenci hominoidů a cerkopiskoidů na 29,6 mil. let (~24-38 mil. let) a divergenci hylobatidů a hominidů na 18,8 mil. let (~16-24 mil. let) (Steiper a Young, 2009). Hominoidi by tedy měli být přítomni již v oligocénu, ale jak je můžeme rozpoznat? Našli jsme je? Identifikace těchto raných fosilních lidoopů je potenciálně ztížena následujícími problémy:
1) Nedostatečný fosilní záznam. V Africe je jen málo oligocenních lokalit, které by reprezentovaly příslušné časové období, a i když existuje několik raně miocenních lokalit, mnoho jejich katarrhinních taxonů je zastoupeno slabě, zejména v postkraniu.
2) Délka doby od vzniku Hominoidea. Bazální příslušníci taxonu se mohou jen málo podobat současným formám, protože si často zachovávají četné primitivní znaky. Například velmi raný lidoop mohl být pronográdní, i když se u posledního společného předka žijících lidoopů vyvinuly adaptace pro vzpřímené držení těla. Očekává se tedy, že potenciálních synapomorfií mezi kmenovými a korunovými hominoidy bude málo.
3) Dlouhá období evoluce v existujících liniích mohla být individuálně jedinečná, takže moderní taxony si zachovaly jen málo určujících synapomorfií, nebo některé z jejich předpokládaných synapomorfií mohou být paralelizmy.
4) Evoluční historie morfologické transformace nemusí být vhodně vyřešena, což vede k rozporuplným fylogenetickým interpretacím.
Srovnávací studie zahrnující jak kmenové, tak korunové katarakty může tyto problémy zmírnit (Zalmout et al., 2010). Například znaky přítomné u pozdně oligocenního nebo raně miocenního druhu, které nejsou primitivně sdíleny s kmenovými katarrhiny a cerkopitékoidy, ale jsou pozorovány u korunových hominoidů, jsou pravděpodobnými hominoidními synapomorfiemi. Například etmofrontální sinus se vyskytuje u afrických lidoopů a chybí u afroarabských kmenových katarrhinů, cercopithekoidů, Pongo a Hylobates (Rossie et al., 2002; Rossie, 2008). Sinus se vyskytuje také u tří domnělých hominoidů z raného miocénu: Proconsul, Morotopithecus a Afropithecus (ibid.). Rozšíření tohoto znaku lze tedy interpretovat jako odvozený znak vyskytující se nejprve u kmenových hominoidů a ztracený u asijských lidoopů (Begun, 1992; Moyà-Solà a Köhler, 1995; alternativní způsoby interpretace tohoto znaku však viz Rossie, 2008). Další lebeční znak, zvýšená příčná robustnost špičáků a současné rozšíření předního patra, má podobné rozšíření (tj, vyskytuje se u současných hominidů (u člověka se druhotně ztratil) i u prokonzula, morotopitéka a afropitéka, ale chybí u kmenových katarrhinů a cerkopitékoidů) a rovněž podporuje interpretaci, že miocenní taxony jsou hominoidi (Begun et al., 1997; Zalmout et al., 2010). Pozdně oligocenní kamoyapithecus je znám pouze z gnatodentálních pozůstatků (Leakey et al., 1995), ale přítomnost příčně rozšířených horních špičáků by mohla signalizovat, že jde rovněž o hominoida (Zalmout et al., 2010). Kromě toho tyto čtyři fosilní rody zahrnují druhy s odhadovanou velikostí těla přesahující 30 kg, což jsou první katarrini, kteří této velikosti dosáhli.
Vzhledem k velkým rozměrům a arboreální nice současných lidoopů (giboni jsou pravděpodobně druhotně zakrslí: Tyler, 1993) může být velká tělesná velikost také kmenovým znakem hominoidů. U korunových hominoidů je tento znak tradičně spojován s hominoidním využíváním kvalitní arboreální potravy, což umožňují lokomoční inovace včetně zvýšené pohyblivosti končetin a schopnosti úchopu (které umožňují rozložení hmotnosti na více opor) a ortografie (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin a Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Důkazy pro tyto postkraniální transformace jsou různě vyjádřeny u prokonsula, afropitheka a morotopitheka. Proconsul je dobře znám z několika částečných koster a byl rekonstruován jako nadpočetný čtyřnožec s pronogradním trupem, ale zdá se, že mu chyběl ocas (Napier a Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). Prokonzul má podle rekonstrukce také zvýšenou schopnost úchopu a větší pohyblivost kloubů, např. loketního, ve srovnání s primitivními katarrhiny a cercopithekoidy, což dokládají znaky, jako je výrazná zona conoidea v distální části pažní kosti (Rose, 1988). Afropithecus je postkraniálně zastoupen méně, ale známými prvky se podobá prokonzulovi (Ward, 1998). Ačkoli spojení mezi kraniálními a postkraniálními vzorky z lokalit v Moroto je předmětem neshod (Senut et al., 2000), postkraniální pozůstatky Morotopithecus se důsledně liší od těch, které jsou připisovány Proconsulovi, a naznačují spíše hominoida s tuhým spodním hřbetem, pohyblivým gleno-humerálním kloubem a femurem přizpůsobeným k úmyslnému šplhání, který více připomíná korunní hominoidy (MacLatchy et al..), 2000).
Kromě větších tělesných rozměrů mají všechny taxony, které lze hodnotit, výrazný pohlavní dimorfismus jak ve velikosti těla, tak v morfologii špičáků (Kelley, 1986; Ruff et al., 1989). Zatímco silný pohlavní dimorfismus je primitivním znakem katarů, u zjevných kmenových hominoidů a moderních lidoopů je dále rozvinut (u člověka je však druhotně redukován).
V chrupu hominoidů existuje jen málo přesvědčivých synapomorfií, pokud vůbec nějaké. Protože si hominoidi zachovávají primitivní morfologii zubů katarrhinů (Kay a Ungar, 1997), zatímco zuby cercopithekoidů sdílejí jasnou synapomorfii molárové bilopodoncie, v minulosti se používaly slabě diagnostické zubní znaky, aby se připustilo, že někteří málo zastoupení oligocénní a miocénní katarrhini mohou být hominoidy, dokud nebudou objeveny kompletnější exempláře (např. Andrews, 1978 a 1992; Stevens et al., 2013). Takoví oligocenní a miocenní katarrini byli neformálně označováni jako „zubaté opice“ (např. Fleagle a Kay, 1987; Cartmill a Smith, 2009) nebo byli zařazeni do taxonomického limbu pomocí označení jako Superfamily incertae sedis (např. Begun, 2007).
Pozoruhodná je myšlenka, že žádný z výše uvedených fosilních taxonů není hominoid a že prvním rozpoznatelným hominoidem je 14 milionů let starý kenyapithecus, který má některé odvozené kraniodentální znaky (např. redukované molární cingulum, vyšší postavení počátku zygomatického oblouku) oproti raně miocenním formám (Harrison, 2010). Pokud by tomu tak bylo, vznikla by mezi odhadovaným vznikem hominoidů a jejich prvním výskytem nevysvětlitelná mezera nejméně 10 mil. let – navzdory robustnímu fosilnímu záznamu po většinu tohoto časového období.
Ačkoli paleoantropologové nejsou schopni s jistotou určit fylogenetické zařazení „zubatých opic“ nebo Kamoyapithecus, existuje široká podpora pro hypotézu, že Proconsul, Afropithecus a Morotopithecus jsou spíše kmenovými hominoidy než kmenovými katarrhiny. Jako okno do rané evoluce hominoidů tyto taxony naznačují následující:
1) Velké tělesné rozměry, zvětšení předního chrupu a jemné změny v zadním chrupu (např. více prodloužené stoličky a změny v umístění stoliček a vývoji cingula) naznačují obsazování zásadně odlišných ekologických nik než u kmenových katarrhinů. Velké špičáky fungují jak při obstarávání potravy, tak při pohlavním výběru, přičemž výrazný dimorfismus naznačuje, že se jednalo o druhý faktor. Velká velikost umožňuje přístup k nové potravě a zlepšuje flexibilitu při hledání potravy (Wheatley, 1987), snižuje riziko predace, podporuje přežívání a ve spojení s faktory, jako je stabilita stanoviště, může souviset s opožděným dospíváním. Analýza vzorce vývoje zubů afropitheků (a možná i prokonzulů) naznačuje opožděné dospívání ve srovnání s ostatními antropoidy (Kelley, 1997, 2002), což z pomalé životní dráhy činí další možnou synapomorfii kmenových hominoidů.
2) Na počátku miocénu existuje mezi pravděpodobnými kmenovými hominoidy s velkým tělem alespoň mírná lokomoční a potravní diverzita, což naznačuje počátky skutečné adaptivní radiace, kdy se současně vyvíjela rostoucí velikost těla, rostoucí lokomoční novost a specializovanější využívání arboreálních zdrojů potravy.
3) V současné době není vyřešeno, které anatomické znaky spojené s ortografií a kloubní pohyblivostí u moderních lidoopů jsou homologní nebo homoplastické (např. Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Tato debata má důsledky pro interpretaci evoluční pozice údajných kmenových hominoidů a pro rekonstrukci vzorce a načasování vzniku adaptací moderních opic.
Například postkraniální podobnosti mezi Morotopithecem a žijícími opicemi lze interpretovat několika způsoby. Pokud žijící opice získaly ortografii paralelní evolucí, pak Morotopithecus představuje nejranější známý případ přechodu, ke kterému v evoluci hominoidů došlo několikrát. To je velmi zajímavé z adaptačního hlediska a naznačuje to, že nějaká základní vývojová nebo genetická tendence (viz níže) řídila evoluční reakci hominoidů stále stejným směrem. Znamenalo by to také, že některé postkraniální znaky je třeba při fylogenetické analýze používat opatrně (Sanders a Bodenbender, 1994). Na druhou stranu, pokud by se tyto lokomoční podobnosti dědily od společného předka, byl by Morotopithecus příbuznější korunovým hominoidům než Proconsul (a případně Afropithecus). Další možností je, že Morotopithecus byl nezávislým „experimentem“ a že společný předek korunních hominoidů byl přesto ortográdní.
4) Jak bylo uvedeno výše, navzdory rozdílům v pohybových specializacích a dlouhým obdobím nezávislé evoluce mají všechny přežívající taxony hominoidů podobný tělesný plán, který je odlišuje od ostatních primátů (s výjimkou některých atletických opic, které mají konvergentně vyvinutý trup a specializaci předních končetin podobnou opicím (Erikson, 1963; Larson, 1998). Základní anatomické základy opičího tělesného plánu vedly k pozoruhodné pohybové rozmanitosti a všestrannosti, která umožňuje velkým tělnatým primátům efektivní pohyb v terénu (pomocí šplhání a zavěšení) i v terestriu (pomocí chůze po kloubech a bipedalismu). Takové lokomoční inovace, spojené s vlastnostmi, jako je ortografie, jsou pravděpodobně základem evolučního „úspěchu“ všech žijících hominoidů a zdá se, že byly do určité míry přítomny i u kmenových hominoidů. Vývojová práce Younga a jeho kolegů (2010) ukázala, že u současných lidoopů je integrace mezi předními a zadními končetinami ve srovnání s kvadrupedními opicemi snížená. Uvolnění tohoto genetického omezení bylo interpretováno tak, že umožnilo končetinám vyvíjet se volněji různými způsoby v reakci na selekční tlaky (ibid.). Odvozené rozdílné používání končetin u morotopitéků (MacLatchy et al., 2013) a mladších miocenních taxonů (např. Moya Sola a Kohler, 1996; Moya Sola et al., 2004; Nakatsukasa a Kunimatsu, 2009) předběžně podporuje domněnku Younga a kolegů (2010), že „vyvinutelnost“ končetin může být ancestrálním znakem hominoidů, ale je zapotřebí více fosilního materiálu a lepšího pochopení genetiky, která je základem tohoto znaku.
Budoucí poznatky o původu a rané evoluci hominoidů vyplynou z pokračujících pokusů o identifikaci a pochopení základů možných hominoidních synapomorfií. To zase bude vyžadovat lepší fosilní záznam kmenových i korunových katarrhinů a podrobnější anatomické, vývojové a genetické studium zkoumaných společných znaků. Integrované studium fosilních hominoidů v jejich paleoenvironmentálním kontextu nám také může pomoci lépe pochopit, proč byly hominoidní rysy selektovány, ať už jednou nebo vícekrát.