Hylobatidit (gibbonit ja siamangit) ovat pienempiä kuin muut elävät apinat, ja ne eroavat toisistaan muillakin tavoin, mikä kuvastaa niiden pitkää evolutiivista eristyneisyyttä hominideista (ihmisapinoista ja ihmisistä), joilla on tuoreempi yhteinen esi-isä (kuva 1). Hominidien (ihmistä lukuun ottamatta) etuhuulet ovat leveämmät kuin hylobatidien (Rae, 1997), niillä on vieläkin lyhyemmät ja jäykemmät alaselät, ja niiltä puuttuu anatomiset erikoistumiset ricochetal brachiationiin (Young, 2003).
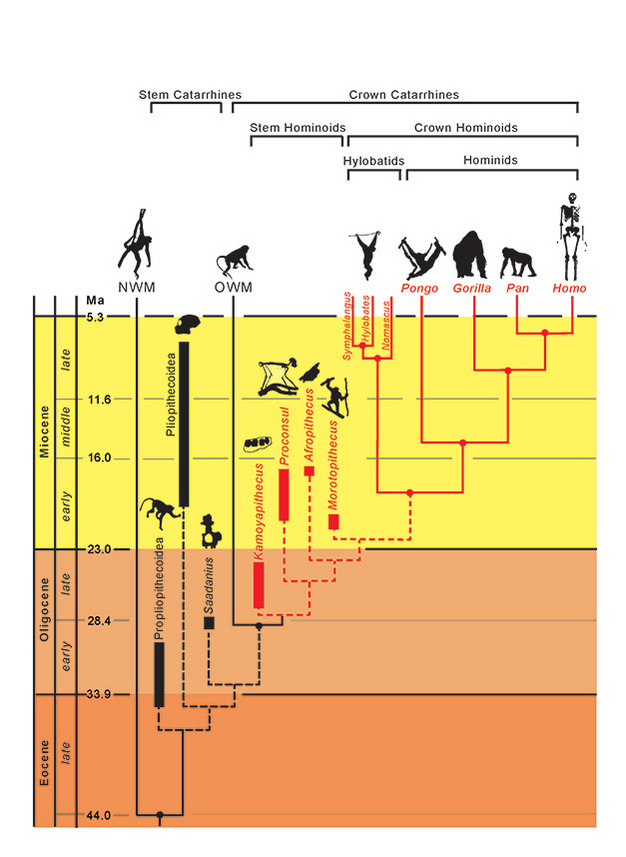
Taksonien fylogeneettiset suhteet on esitetty yhtenäisillä (elossa olevat) ja katkoviivoilla (kuolleet). Geologinen aika on esitetty pystyakselilla. Taksonien haarautumistapahtumien ajoitus on vain likimääräinen, ja se on johdettu molekyyli- ja fossiilitietojen yhdistelmästä (Steiper et al., 2004; MacLatchy et al., 2006; Steiper ja Young, 2008 ja 2009; Chan et al., 2010; Stevens et al., 2013). Fossiilisten taksonien fylogeneettiset sijoitukset ovat hypoteettisia, kuten tekstissä käsitellään. Afropithecuksen ja Morotopithecuksen asema voi olla päinvastainen, tai ne voivat olla sisartaksoneita. NMW viittaa Uuden maailman apinoihin ja OWM viittaa Vanhan maailman apinoihin, kun taas Propliopithecoidea, Pliopithecoidea ja Saadanius viittaavat Afro-Arabiasta peräisin oleviin kantakatarriineihin, joilta puuttuu kruunukatarriinien synapomorfiat.
Kun otetaan huomioon nämä adaptiiviset samankaltaisuudet elävien hominoidien keskuudessa, on mielenkiintoista tutkia, mitkä (jos mitkään) piirteet luonnehtivat hominoidien alkuperäistä erilaistumista muista kädellisistä katarriineista, erityisesti cercopithecoideista (vanhan maailman apinoista). Yhdistettyjen morfologisten ja geneettisten todisteiden mukaan hominoidien ja serkopithekoidien välinen eroavaisuus ajoittuu 29,6 Ma:n (~24-38 Ma) ja hylobatidien ja hominidien välinen eroavaisuus 18,8 Ma:n (~16-24 Ma) aikaan (Steiper ja Young, 2009). Hominoideja pitäisi siis esiintyä joligoseeniin mennessä, mutta miten voimme tunnistaa ne? Olemmeko löytäneet niitä? Näiden varhaisten fossiilisten apinoiden tunnistamista vaikeuttavat mahdollisesti seuraavat ongelmat:
1) Puutteelliset fossiilitiedot. Oligoseenin aikaisia, sopivaa ajanjaksoa edustavia löytöpaikkoja on Afrikassa vain vähän, ja vaikka varhaisen mioseenin aikaisia löytöpaikkoja on useita, monet niiden katarriiniset taksonit ovat heikosti edustettuina, erityisesti postcraniumissa.
2) Hominoidean synnystä kulunut aika. Taksonin perusjäsenet saattavat muistuttaa vain vähän nykyisiä muotoja, koska niissä on usein säilynyt lukuisia primitiivisiä piirteitä. Esimerkiksi hyvin varhainen apina on voinut olla pronograde, vaikka elävien apinoiden viimeisellä yhteisellä esi-isällä olisi kehittynyt sopeutumia pystyasentoon. Mahdollisia synapomorfioita kanto- ja kruunuhominoidien välillä odotetaan näin ollen olevan vain vähän.
3) Pitkät evoluutiojaksot elossa olevissa sukulinjoissa ovat voineet olla yksilöllisesti ainutlaatuisia siten, että nykyaikaisilla taksoneilla on säilynyt vain vähän määritteleviä synapomorfioita, tai jotkut niiden oletetuista synapomorfioista voivat olla rinnakkaisuuksia.
4) Morfologisen muodonmuutoksen evoluutiohistoriaa ei ehkä ole asianmukaisesti ratkaistu, mikä johtaa ristiriitaisiin fylogeneettisiin tulkintoihin.
Vertaileva tutkimus, jossa on mukana sekä varsi- että kruunukatarriineja, voi lieventää näitä ongelmia (Zalmout ym., 2010). Esimerkiksi myöhäisligoseeni- tai varhaismioseenilajissa esiintyvät piirteet, joita ei jaeta primitiivisesti kantakatarriinien ja cercopithecoidien kanssa, mutta joita havaitaan kruunuhominoideilla, ovat uskottavia hominoidien synapomorfioita. Esimerkiksi ethmo-frontal sinus esiintyy afrikkalaisilla apinoilla, ja se puuttuu afroarabialaisilta kantakatarriineilta, cercopithecoideilta, Pongolta ja Hylobatesilta (Rossie et al., 2002; Rossie, 2008). Sinus esiintyy myös kolmella varhaismioseenisen ajan oletetulla hominoidilla: Proconsul, Morotopithecus ja Afropithecus (ibid.). Tämän piirteen levinneisyys voidaan siis tulkita johdetuksi piirteeksi, joka esiintyi ensin kantahominoideilla ja hävisi aasialaisista apinoista (Begun, 1992; Moyà-Solà ja Köhler, 1995; ks. kuitenkin Rossie, 2008, vaihtoehtoisista tavoista tulkita tämä piirre). Toisella kallonpiirteellä, kulmahampaiden lisääntyneellä poikkileikkauksen kestävyydellä ja samanaikaisella etummaisen suulakihampaan laajenemisella, on samanlainen levinneisyys (ts, sitä esiintyy nykyisillä hominideilla (hävinnyt toissijaisesti ihmisillä) sekä Proconsulilla, Morotopithecuksella ja Afropithecuksella, mutta sitä ei esiinny kantakatarriineilla ja cercopithecoideilla), ja se tukee myös tulkintaa, jonka mukaan mioseeniset taksonit ovat hominoideja (Begun ym., 1997; Zalmout ym., 2010). Oligoseenin lopun Kamoyapithecus tunnetaan vain gnathodentaalisten jäänteiden perusteella (Leakey ym., 1995), mutta poikkileikkaukseltaan laajentuneiden yläetuhampaiden läsnäolo voisi olla merkki siitä, että myös se on hominoidi (Zalmout ym., 2010). Lisäksi näihin neljään fossiiliseen sukuun kuuluu lajeja, joiden arvioitu ruumiinkoko on yli 30 kg, ja ne ovat ensimmäiset tämän koon saavuttaneet katarriinit.
Kun otetaan huomioon nykyisten apinoiden suuri koko ja arboreaalinen niche (gibbonit ovat luultavasti toissijaisesti kääpiökasvuisia: Tyler, 1993), suuri ruumiinkoko voi olla myös varsihominoidien ominaisuus. Kruunuhominoideilla tämä ominaisuus on perinteisesti liitetty hominoidien korkealaatuisen arboreaalisen ravinnon hyödyntämiseen, jonka mahdollistavat lokomotoriset innovaatiot, joihin kuuluvat raajojen lisääntynyt liikkuvuus ja tarttumiskyky (jotka mahdollistavat painon jakautumisen useille kannattimille) sekä ortogradia (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin ja Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Todisteet näistä kallon jälkeisistä muutoksista ovat vaihtelevasti ilmaistu Proconsul-, Afropithecus- ja Morotopithecus-lajeissa. Proconsul tunnetaan hyvin useista osittaisista luurangoista, ja se on rekonstruoitu ylävartaloiseksi nelijalkaiseksi, jolla oli pronogradi vartalo, mutta siltä näyttää puuttuneen häntä (Napier ja Davis, 1959; Beard ym., 1986; Ward ym., 1991; Ward, 1998). Proconsulilla on myös rekonstruoitu olevan parempi tarttumiskyky ja suurempi liikkuvuus nivelissä, kuten kyynärnivelessä, suhteessa alkukantaisiin katarriineihin ja cercopithecoideihin, mistä ovat osoituksena sellaiset piirteet kuin erillinen zona conoidea distaalisessa humeruksessa (Rose, 1988). Afropithecus on vähemmän hyvin edustettuna postcraniaalisesti, mutta muistuttaa Proconsulia tunnetuilta osiltaan (Ward, 1998). Vaikka Moroto-löydösten kallo- ja kallonjälkeisten näytteiden välisestä yhteydestä ollaan eri mieltä (Senut ym., 2000), Morotopithecuksen kallonjälkeiset jäännökset poikkeavat johdonmukaisesti Proconsulille liitetyistä jäännöksistä ja viittaavat sen sijaan hominoidiin, jolla on jäykkä alaselkä, liikkuva nilkkanivel ja tarkoitukselliseen kiipeilyyn sopeutunut reisiluu, mikä muistuttaa enemmän kruunuhominoideja (MacLatchy ym., 2000), 2000).
Suurempien ruumiinkokojen lisäksi kaikilla arvioitavissa olevilla taksoneilla on selvä sukupuolidimorfismi sekä ruumiinkoon että koiraiden morfologian suhteen (Kelley, 1986; Ruff ym., 1989). Vaikka voimakas seksuaalinen dimorfismi on primitiivinen katarriinien piirre, se on kehittyneempi ilmeisissä kantahominoideissa ja nykyaikaisissa ihmisapinoissa (mutta toissijaisesti vähentynyt ihmisillä).
Hominoidien hampaistossa on vain vähän, jos lainkaan, pakottavia synapomorfioita. Koska hominoidit säilyttävät primitiivisen katarriinihampaiden morfologian (Kay ja Ungar, 1997), kun taas sercopithecoidien hampaiden selkeä synapomorfia on molaarinen bilofodontia, on ollut tapana käyttää heikosti diagnostisia hammasominaisuuksia sen tunnustamiseksi, että jotkin heikosti edustetut oligoseeni- ja miooseenikauden katarriinihampaat saattaisivat olla hominoideja, kunnes täydellisempiä yksilöitä löydetään (esim. Andrews, 1978 ja 1992; Stevens ym., 2013). Tällaisia oligoseeni- ja miocene-katarriineja on kutsuttu epävirallisesti ”hammasapinoiksi” (esim. Fleagle ja Kay, 1987; Cartmill ja Smith, 2009), tai ne on sijoitettu taksonomiseen limboon käyttämällä nimitystä Superfamily incertae sedis (esim. Begun, 2007).
Huomattakoon, että on esitetty ajatus, että yksikään edellä käsitellyistä fossiilisista taksoneista ei ole hominoidi ja että ensimmäinen tunnistettava hominoidi on 14 miljoonaa vuotta vanha Kenyapithecus, jolla on joitain johdettuja kraniodentaalisia ominaisuuksia (esim. pienentynyt molaarinen cingulum, poskihammaskaaren korkeampi syntyasento) suhteessa varhaiseen mioseeniin kuuluviin muotoihin (Harrison, 2010). Jos näin on, hominoidien arvioidun alkuperän ja niiden ensiesiintymisen välille jäisi selittämätön vähintään 10 ma:n aukko – huolimatta vankasta fossiiliaineistosta suurelta osin kyseiseltä ajanjaksolta.
Vaikka paleoantropologit eivät pysty määrittelemään ”hammasapinoiden” tai Kamoyapithecuksen fylogeneettistä sijoittumista luotettavasti, on olemassa laajaa tukea hypoteesille, jonka mukaan Proconsul, Afropithecus ja Morotopithecus ovat pikemminkin kantahominoideja kuin kantakatarriineja. Ikkunana varhaiseen hominoidien evoluutioon nämä taksonit viittaavat seuraaviin seikkoihin:
1) Suuri ruumiinkoko, etuhampaiston laajentuminen ja hienovaraiset muutokset takahampaistossa (esim. pidemmät poskihampaat ja muutokset poskihampaiden kärkien sijoittelussa ja kitalakeen kehityksessä) viittaavat siihen, että ne ovat miehittäneet perustavanlaatuisesti erilaisia ekologisia markkinarakoja kuin kantakatarriinit. Suuret kulmahampaat toimivat sekä ravinnon hankinnassa että sukupuolisessa valinnassa, ja voimakas dimorfismi viittaa siihen, että jälkimmäinen oli tekijä. Suuri koko mahdollistaa uusien ravintokohteiden saannin ja parantaa ravinnonhankinnan joustavuutta (Wheatley, 1987), vähentää saalistusriskiä, edistää eloonjäämistä, ja yhdessä elinympäristön vakauden kaltaisten tekijöiden kanssa se voi olla yhteydessä myöhäisempään aikuistumiseen. Afropithecuksen (ja mahdollisesti Proconsulin) hampaiden kehitysmallin analyysi viittaa viivästyneeseen kypsymiseen suhteessa muihin antropoideihin (Kelley, 1997, 2002), mikä tekee hitaasta elämänhistoriasta toisen mahdollisen kantahominoidien synapomorfian.
2) Varhaismioeniin mennessä suurikokoisten todennäköisten kantohominoidien joukossa on ainakin vaatimatonta lokomotorista ja ravitsemuksellista monimuotoisuutta, mikä viittaa todellisen adaptiivisen säteilyn alkuun, jossa kasvava ruumiinkoko, lisääntyvä lokomotorinen uutuus ja arboreaalisten ravinnonlähteiden erikoistuneempi hyväksikäyttö kehittyvät samaan aikaan.
3) Tällä hetkellä ei ole ratkaistavissa, mitkä ortogradiaan ja nivelten liikkuvuuteen liittyvät anatomiset piirteet nykyaikaisilla apinoilla ovat homologisia tai homoplastisia (esim. Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Tällä keskustelulla on vaikutuksia väitettyjen kantahominoidien evolutiivisen aseman tulkintaan ja nykyapinoiden sopeutumismallien syntymismallin ja -ajankohdan rekonstruointiin.
Esimerkiksi Morotopithecuksen ja elävien apinoiden kallonjälkeisiä yhtäläisyyksiä voidaan tulkita monella tavalla. Jos elävät apinat saivat ortogradiansa rinnakkaisen evoluution kautta, Morotopithecus edustaa varhaisinta tunnettua tapausta siirtymästä, joka tapahtui useita kertoja hominoidien evoluutiossa. Tämä on erittäin kiinnostavaa sopeutumisen kannalta, sillä se viittaa siihen, että jokin taustalla oleva kehityspiirre tai geneettinen taipumus (ks. jäljempänä) ohjasi hominoidien evoluutiota samaan suuntaan. Se merkitsisi myös sitä, että tiettyjä kallon jälkeisiä piirteitä on käytettävä varovaisesti fylogeneettisessä analyysissä (Sanders ja Bodenbender, 1994). Toisaalta, jos nämä liikuntaelinten samankaltaisuudet olisivat periytyneet yhteisestä esi-isästä, Morotopithecus olisi läheisempää sukua kruunuhominoideille kuin Proconsul (ja mahdollisesti Afropithecus). Toinen mahdollisuus on, että Morotopithecus oli itsenäinen ”kokeilu” ja että kruunuhominoidien yhteinen esi-isä oli kuitenkin ortogradinen.
4) Kuten edellä käsiteltiin, huolimatta eroista liikuntaelinten erikoistumisessa ja pitkistä itsenäisen evoluution ajanjaksoista, kaikilla elossa olevilla hominoidien taksoneilla on samankaltainen ruumiinrakenne, joka erottaa ne muista kädellisistä (lukuun ottamatta eräitä ateljeeapinoita, joilla on konvergentisti kehittynyt apinoiden kaltainen torso ja eturaajojen erikoistuminen (Erikson, 1963; Larson, 1998). Apinoiden ruumiinrakenteen perusanatomia on johtanut huomattavaan liikuntaelimistön monimuotoisuuteen ja monipuolisuuteen, minkä ansiosta suurikokoiset kädelliset voivat liikkua tehokkaasti arboreaalisesti (kiipeilemällä ja ripustautumalla) ja terrestrisesti (kävelemällä rystysillä ja kaksijalkaisesti). Tällainen ortogradian kaltaisiin ominaisuuksiin sidoksissa oleva liikuntainnovaatio on todennäköisesti kaikkien elävien hominoidien evolutiivisen ”menestyksen” perusta, ja se näyttää olleen jossain määrin läsnä myös kantahominoideilla. Youngin ja kollegoiden (2010) kehitystyö on osoittanut, että nykyisten apinoiden etu- ja takaraajojen välinen integraatio on heikentynyt nelijalkaisiin apinoihin verrattuna. Tämän geneettisen rajoituksen höllentymisen on tulkittu antaneen raajojen kehittyä vapaammin eri tavoin vastauksena valintapaineisiin (ibid.). Morotopithecuksen (MacLatchy ym., 2013) ja nuorempien mioseenitaksonien (esim. Moya Sola ja Kohler, 1996; Moya Sola ym., 1996; Moya Sola ym, 2004; Nakatsukasa ja Kunimatsu, 2009) tukee alustavasti Youngin ja kollegoiden (2010) olettamusta, jonka mukaan raajojen ”evolvoituvuus” voi olla hominoidien esi-isien ominaisuus, mutta tarvitaan lisää fossiiliaineistoa ja parempaa ymmärrystä tämän ominaisuuden taustalla olevasta genetiikasta.
Tulevaisuuden oivallus hominoidien alkuperästä ja varhaisesta evoluutiosta syntyy, kun jatketaan pyrkimyksiä identifioida ja ymmärtää mahdollisten hominoidien synapomorfioiden perustaa. Tämä puolestaan edellyttää parempia fossiiliaineistoja sekä varsi- että kruunukatarriineista ja yksityiskohtaisempaa anatomista, kehityksellistä ja geneettistä tutkimusta tarkastelun kohteena olevista yhteisistä piirteistä. Fossiilisten hominoidien kokonaisvaltainen tutkiminen niiden paleoympäristökontekstissa voi myös auttaa meitä ymmärtämään paremmin, miksi hominoidien piirteet valikoituivat joko kerran tai useita kertoja.