T.H. Huxley a écrit que regarder les autres singes, c’est voir « des copies floues de soi-même » (1863 pg. 73). L’étroite relation physique entre les singes, ou hominoïdes, avait déjà été reconnue un siècle plus tôt par Carolus Linnaeus (1759). Les hominoïdes vivants sont unis par des caractéristiques liées à l’orthogradie habituelle et aux comportements en dessous de la branche : un large torse avec des articulations d’épaule très espacées, un dos rigide, de longs membres antérieurs, des articulations de membres mobiles, une forte capacité de préhension et l’absence de queue (Huxley, 1863 ; Harrison, 1987 & 1991 ; Shoshani et al., 1996). Les humains sont dépourvus de plusieurs de ces caractéristiques, comme des bras relativement longs, en raison d’une adaptation secondaire à la bipédie. Des attributs crâniens unissent également les hominoïdes, notamment des palais antérieurs larges et des cerveaux relativement grands (Jerison, 1973 ; Rae, 1997). Les hominoïdes ont également une durée de vie prolongée avec une maturation retardée (Kelley, 1997), et occupent des niches écologiques similaires – toutes les espèces, sauf les humains, habitent des forêts tropicales et dépendent considérablement des fruits mûrs et des feuilles pour se nourrir (Fleagle, 2013).
Les hylobatides (gibbons et siamangs) sont plus petits que les autres grands singes vivants et sont distincts à d’autres égards, reflétant leur longue période d’isolement évolutif par rapport aux hominidés (grands singes et humains), qui partagent un ancêtre commun plus récent (Figure 1). Les hominidés (à l’exception des humains) ont des palais antérieurs élargis au-delà de ceux des hylobatidés (Rae, 1997), possèdent des bas de dos encore plus courts et plus rigides, et manquent de spécialisations anatomiques pour la brachiation ricochet (Young, 2003).
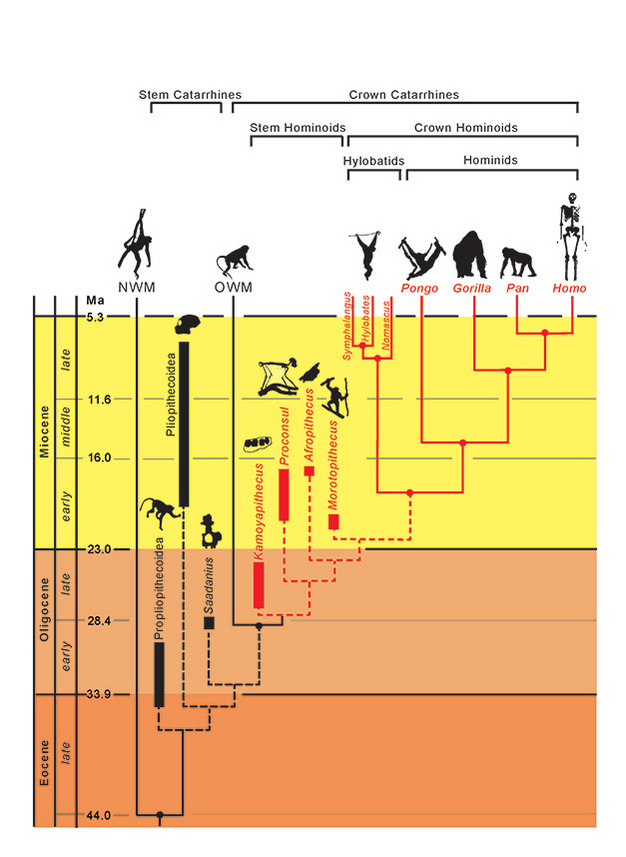
Les relations phylogénétiques des taxons sont représentées par des lignes pleines (éteintes) et en pointillés (éteintes). Le temps géologique est représenté sur l’axe vertical. La chronologie des embranchements des taxons est approximative et déduite d’une combinaison de données moléculaires et fossiles (Steiper et al., 2004 ; MacLatchy et al., 2006 ; Steiper et Young, 2008 et 2009 ; Chan et al., 2010 ; Stevens et al., 2013). Les placements phylogénétiques des taxons fossiles sont hypothétiques, comme discuté dans le texte. La position d’Afropithecus et de Morotopithecus pourrait être inversée, ou les deux pourraient être des taxons frères. NMW se réfère aux singes du Nouveau Monde et OWM aux singes de l’Ancien Monde, tandis que Propliopithecoidea, Pliopithecoidea et Saadanius se réfèrent aux catarrhines à tige d’Afro-Arabie dépourvus de synapomorphies de catarrhines à couronne.
Compte tenu de ces similitudes adaptatives parmi les hominoïdes vivants, il est intéressant de rechercher lesquelles (le cas échéant) ont caractérisé la divergence initiale des hominoïdes par rapport aux autres primates catarrhines, notamment les cercopithécoïdes (singes de l’Ancien Monde). Des preuves morphologiques et génétiques combinées situent la divergence entre les hominoïdes et les cercopithécoïdes à 29,6 Ma (~24-38 Ma), et la divergence entre les hylobatides et les hominidés à 18,8 Ma (~16-24 Ma) (Steiper et Young, 2009). Les hominoïdes devraient donc être présents dès l’Oligocène, mais comment les reconnaître ? Les avons-nous trouvés ? L’identification de ces premiers singes fossiles est potentiellement entravée par les problèmes suivants :
1) Un registre fossile déficient. Il y a peu de sites oligocènes en Afrique qui représentent la période appropriée, et bien qu’il y ait plusieurs sites du début du Miocène, beaucoup de leurs taxons catarrhines sont mal représentés, en particulier dans le postcranium.
2) Le temps écoulé depuis l’origine des Hominoidea. Les membres basaux d’un taxon peuvent ressembler peu aux formes existantes car ils conservent souvent de nombreux caractères primitifs. Par exemple, un singe très ancien pourrait avoir été pronograde même si le dernier ancêtre commun des singes vivants avait développé des adaptations pour une posture droite. Les synapomorphies potentielles entre les hominoïdes à tige et à couronne devraient donc être peu nombreuses.
3) Les longues périodes d’évolution des lignées existantes peuvent avoir été individuellement uniques de sorte que les taxons modernes conservent peu de synapomorphies déterminantes, ou certaines de leurs synapomorphies présumées peuvent être des parallélismes.
4) L’histoire évolutive de la transformation morphologique peut ne pas être résolue de manière appropriée, ce qui entraîne des interprétations phylogénétiques contradictoires.
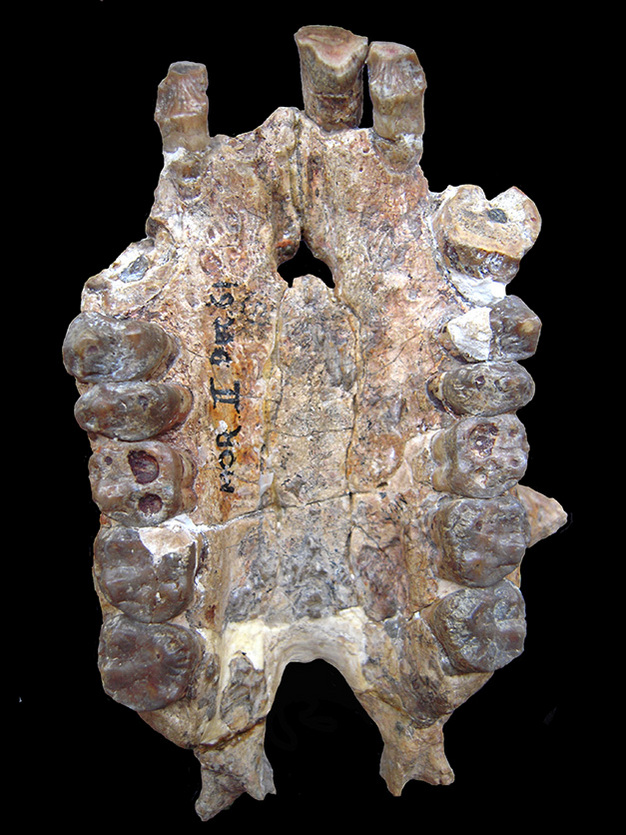
Une étude comparative impliquant à la fois des catarrhines à tige et à couronne peut atténuer ces problèmes (Zalmout et al., 2010). Par exemple, les caractéristiques présentes chez une espèce de la fin de l’Oligocène ou du début du Miocène, qui ne sont pas partagées primitivement avec les catarrhines à tige et les cercopithécoïdes, mais qui sont observées chez les hominoïdes à couronne, sont des synapomorphies hominoïdes plausibles. Par exemple, le sinus ethmo-frontal est présent chez les singes africains et est absent chez les catarrhines à tige afro-arabes, les cercopithécoïdes, Pongo et Hylobates (Rossie et al., 2002 ; Rossie, 2008). Le sinus est également présent chez trois hominoïdes putatifs du Miocène précoce : Proconsul, Morotopithecus et Afropithecus (ibid.). La distribution de ce trait peut donc être interprétée comme un caractère dérivé apparaissant d’abord chez les hominoïdes souches, et perdu chez les singes asiatiques (Begun, 1992 ; Moyà-Solà et Köhler, 1995 ; mais voir Rossie, 2008, pour d’autres façons d’interpréter ce caractère). Une autre caractéristique crânienne, l’augmentation de la robustesse de la section transversale des canines, et l’expansion concomitante du palais antérieur, a une distribution similaire (c’est-à-dire, elle est présente chez les hominidés existants (perdue secondairement chez l’homme) ainsi que chez Proconsul, Morotopithecus et Afropithecus, mais est absente chez les catarrhines et les cercopithécoïdes), et soutient également l’interprétation selon laquelle les taxons miocènes sont des hominoïdes (Begun et al., 1997 ; Zalmout et al., 2010). Le Kamoyapithecus de l’Oligocène tardif n’est connu que par des restes gnathodentaires (Leakey et al., 1995), mais la présence de canines supérieures élargies en coupe pourrait indiquer qu’il s’agit également d’un hominoïde (Zalmout et al., 2010). En outre, ces quatre genres fossiles comprennent des espèces dont la taille corporelle estimée dépasse 30 kg, les premiers catarrhines à atteindre cette taille.
Compte tenu de la grande taille et de la niche arboricole des grands singes existants (les gibbons sont probablement secondairement nains : Tyler, 1993), la grande taille du corps peut également être une caractéristique des hominoïdes souches. Chez les hominoïdes de la couronne, ce trait est traditionnellement lié à l’exploitation par les hominoïdes d’aliments arboricoles de haute qualité, permise par des innovations locomotrices, notamment une mobilité accrue des membres et une capacité de préhension (qui permettent de répartir le poids sur plusieurs supports), ainsi que l’orthogradie (Avis, 1962 ; Napier, 1967 ; Tuttle, 1975 ; Fleagle, 1976 ; Cachel, 1979 ; Temerin et Cant, 1983 ; Cant, 1987 ; Wheatley, 1987 ; Kelley, 1997 ; MacLatchy, 2004).
Les preuves de ces transformations postcrâniennes sont exprimées de manière variable chez Proconsul, Afropithecus et Morotopithecus. Proconsul est bien connu grâce à plusieurs squelettes partiels et a été reconstitué comme un quadrupède supra-branché, avec un tronc pronograde, mais semble ne pas avoir eu de queue (Napier et Davis, 1959 ; Beard et al., 1986 ; Ward et al., 1991 ; Ward, 1998). Proconsul est également reconstitué comme ayant une meilleure capacité de préhension et une plus grande mobilité des articulations telles que le coude, par rapport aux catarrhines et aux cercopithèques primitifs, comme en témoignent des caractéristiques telles qu’une zone conoidea distincte dans l’humérus distal (Rose, 1988). Afropithecus est moins bien représenté au niveau postcrânien, mais ressemble à Proconsul par des éléments connus (Ward, 1998). Bien que l’association entre les échantillons crâniens et postcrâniens des localités de Moroto fasse l’objet de désaccords (Senut et al., 2000), les restes postcrâniens de Morotopithecus diffèrent systématiquement de ceux attribués à Proconsul, suggérant plutôt un hominoïde au bas du dos raide, à l’articulation gléno-humérale mobile et au fémur adapté à l’escalade délibérée, rappelant davantage les hominoïdes de la couronne (MacLatchy et al, 2000).
En plus des tailles corporelles plus importantes, tous les taxons qui peuvent être évalués présentent un dimorphisme sexuel prononcé tant au niveau de la taille corporelle que de la morphologie canine (Kelley, 1986 ; Ruff et al., 1989). Alors qu’un fort dimorphisme sexuel est une caractéristique catarrhine primitive, il est plus élaboré chez les hominoïdes souches apparents et les grands singes modernes (mais secondairement réduit chez les humains).
Il y a peu, voire pas, de synapomorphies convaincantes dans la dentition des hominoïdes. Parce que les hominoïdes conservent la morphologie primitive des dents des catarrhines (Kay et Ungar, 1997), tandis que les dents des cercopithécoïdes partagent la synapomorphie claire et nette de la bilophodontie molaire, il y a une histoire d’utilisation de caractéristiques dentaires faiblement diagnostiques pour reconnaître que certains catarrhines oligocènes et miocènes mal représentés pourraient être des hominoïdes, en attendant la découverte de spécimens plus complets (par exemple Andrews, 1978 et 1992 ; Stevens et al., 2013). Ces catarrhines de l’Oligocène et du Miocène ont été désignés de manière informelle comme des » singes dentaires » (par exemple Fleagle et Kay, 1987 ; Cartmill et Smith, 2009), ou placés dans les limbes taxonomiques en utilisant une désignation telle que Superfamily incertae sedis (par exemple Begun, 2007).
Notamment, l’idée a été avancée qu’aucun des taxons fossiles discutés ci-dessus n’est un hominoïde, et que le premier hominoïde reconnaissable est le Kenyapithecus, vieux de 14 millions d’années, qui présente certains attributs craniodentaux dérivés (par exemple, un cingulum molaire réduit, une position d’origine de l’arc zygomatique plus élevée) par rapport aux formes du Miocène précoce (Harrison, 2010). Si c’est le cas, il y aurait un écart inexplicable d’au moins 10 Ma – malgré un registre fossile robuste pendant une grande partie de cette période – entre l’origine estimée des hominoïdes et leur première apparition.
Bien que les paléoanthropologues ne soient pas en mesure de déterminer le placement phylogénétique des » singes dentaires » ou de Kamoyapithecus avec confiance, il existe un large soutien pour l’hypothèse selon laquelle Proconsul, Afropithecus et Morotopithecus sont des hominoïdes souches, plutôt que des catarrhines souches. En tant que fenêtre sur l’évolution précoce des hominoïdes, ces taxons suggèrent ce qui suit :
1) La grande taille du corps, l’élargissement de la dentition antérieure et les changements subtils dans la dentition postérieure (par exemple, des molaires plus allongées et des changements dans le placement des cuspides des molaires et le développement du cingulum) suggèrent l’occupation de niches écologiques fondamentalement différentes de celles des catarrhines souches. Les grandes canines servent à la fois à l’acquisition de nourriture et à la sélection sexuelle, avec un dimorphisme prononcé suggérant que ce dernier facteur a joué un rôle. La grande taille permet l’accès à de nouveaux aliments et améliore la flexibilité de la recherche de nourriture (Wheatley, 1987), diminue les risques de prédation, favorise la survie et, associée à des facteurs tels que la stabilité de l’habitat, peut être liée à une maturation retardée. L’analyse du modèle de développement dentaire d’Afropithecus (et peut-être de Proconsul) suggère une maturation retardée par rapport aux autres anthropoïdes (Kelley, 1997, 2002), faisant de l’histoire de vie lente un autre synapomorphie possible des hominoïdes souches.
2) Au début du Miocène, il existe une diversité locomotrice et alimentaire au moins modeste parmi les hominoïdes souches probables à grand corps, indiquant les débuts d’une véritable radiation adaptative, avec une taille corporelle croissante, une nouveauté locomotrice croissante et une exploitation plus spécialisée des sources alimentaires arboricoles évoluant simultanément.
3) Il n’est pas actuellement possible de résoudre quelles caractéristiques anatomiques associées à l’orthogradie et à la mobilité articulaire chez les grands singes modernes sont homologues ou homoplastiques (par exemple, Harrison, 1991 ; Larson, 1998 ; Young, 2003 ; MacLatchy, 2004 & 2010 ; Lovejoy et al., 2009). Ce débat a des implications pour l’interprétation de la position évolutive des hominoïdes souches présumés, et pour la reconstitution du modèle et du moment de l’émergence des adaptations des singes modernes.
Par exemple, les similitudes postcrâniennes entre Morotopithecus et les singes vivants pourraient être interprétées de plusieurs façons. Si les grands singes vivants ont acquis leur orthogénie par une évolution parallèle, alors Morotopithecus représente le premier exemple connu d’une transition qui s’est produite plusieurs fois dans l’évolution des hominoïdes. Ceci est d’un grand intérêt adaptatif, suggérant qu’une certaine propension développementale ou génétique sous-jacente (voir ci-dessous) a continué à orienter la réponse évolutive des hominoïdes dans la même direction. Cela signifierait également que certains traits postcrâniens doivent être utilisés avec précaution dans les analyses phylogénétiques (Sanders et Bodenbender, 1994). D’autre part, si ces similitudes locomotrices étaient héritées d’un ancêtre commun, Morotopithecus serait plus proche des hominoïdes de la couronne que Proconsul (et peut-être Afropithecus). Une autre possibilité encore est que Morotopithecus était une « expérience » indépendante, et que l’ancêtre commun des hominoïdes à couronne était néanmoins orthograde.
4) Comme discuté ci-dessus, malgré des différences dans les spécialisations locomotrices et de longues périodes d’évolution indépendante, tous les taxons d’hominoïdes survivants partagent un plan corporel similaire qui les différencie des autres primates (à l’exception de certains singes atèles, qui ont évolué de manière convergente vers des torses et des spécialisations des membres antérieurs similaires à ceux des grands singes (Erikson, 1963 ; Larson, 1998). Les fondements anatomiques du plan corporel des grands singes ont conduit à une diversité et une polyvalence locomotrices remarquables, permettant aux grands primates de se déplacer efficacement dans les arbres (en grimpant et en se suspendant) et sur la terre ferme (en marchant avec les articulations et en étant bipède). Cette innovation locomotrice, liée à des caractéristiques telles que l’orthogradie, est probablement à la base du « succès » évolutif de tous les hominoïdes vivants, et semble avoir été présente à un certain degré chez les hominoïdes souches. Les travaux de Young et de ses collègues (2010) sur le développement ont montré que les grands singes existants ont une intégration réduite entre les membres antérieurs et postérieurs par rapport aux singes quadrupèdes. La relaxation de cette contrainte génétique a été interprétée comme ayant permis aux membres d’évoluer plus librement de différentes manières en réponse aux pressions de sélection (ibid.). L’utilisation différentielle présumée des membres chez Morotopithecus (MacLatchy et al., 2013) et chez des taxons miocènes plus jeunes (par exemple Moya Sola et Kohler, 1996 ; Moya Sola et al, 2004 ; Nakatsukasa et Kunimatsu, 2009) soutient provisoirement la supposition de Young et de ses collègues (2010) selon laquelle l' » évolutivité » des membres pourrait être un trait hominoïde ancestral, mais davantage de matériel fossile et une meilleure compréhension de la génétique sous-jacente à ce trait sont nécessaires.
La compréhension future des origines des hominoïdes et de leur évolution précoce émergera des tentatives continues d’identifier et de comprendre la base des synapomorphies hominoïdes possibles. Ceci, à son tour, nécessitera une amélioration des archives fossiles pour les catarrhines à tige et à couronne, ainsi qu’une étude anatomique, développementale et génétique plus détaillée des caractères communs examinés. L’étude intégrée des hominoïdes fossiles dans leur contexte paléoenvironnemental peut également nous aider à mieux comprendre pourquoi les caractéristiques des hominoïdes ont été sélectionnées, une ou plusieurs fois.