T.H. Huxley ha scritto che guardare le altre scimmie è vedere “copie sfocate di se stessi” (1863 pag. 73). La stretta relazione fisica tra le scimmie, o ominoidi, era già stata riconosciuta un secolo prima da Carolus Linnaeus (1759). Gli ominoidi viventi sono accomunati da caratteristiche legate all’ortogonalità abituale e ai comportamenti al di sotto del tronco: torsi ampi con articolazioni delle spalle molto distanziate, schiene rigide, arti anteriori lunghi, articolazioni degli arti mobili, forte capacità di afferrare e assenza di coda (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Gli esseri umani mancano di molte di queste caratteristiche, come le braccia relativamente lunghe, come risultato di un adattamento secondario al bipedalismo. Anche gli attributi cranici accomunano gli ominoidi, tra cui ampi palati anteriori e cervelli relativamente grandi (Jerison, 1973; Rae, 1997). Gli ominidi hanno anche una durata di vita estesa con maturazione ritardata (Kelley, 1997), e occupano nicchie ecologiche simili – tutte le specie tranne gli esseri umani abitano le foreste tropicali e si basano significativamente su frutta matura e foglie per il cibo (Fleagle, 2013).
Gli ominidi (gibboni e siamang) sono più piccoli delle altre scimmie viventi e sono distinti in altri modi, riflettendo il loro lungo periodo di isolamento evolutivo dagli ominidi (grandi scimmie e uomini), che condividono un antenato comune più recente (Figura 1). Gli ominidi (tranne gli umani) hanno palati anteriori che sono allargati oltre quelli degli hylobatidi (Rae, 1997), possiedono dorsi inferiori ancora più corti e più rigidi, e mancano di specializzazioni anatomiche per la brachiazione ricochetale (Young, 2003).
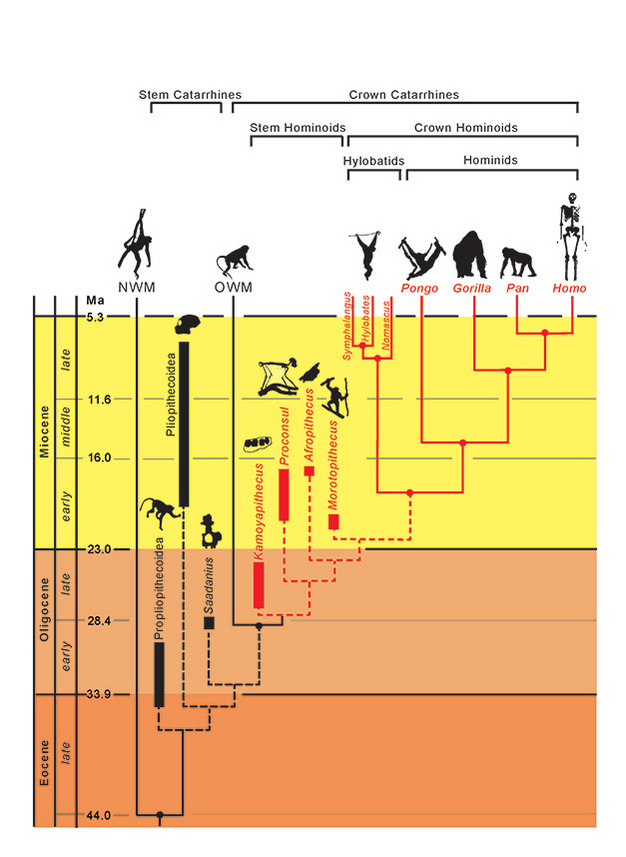
Le relazioni filogenetiche dei taxa sono rappresentate da linee solide (estinti) e tratteggiate (estinti). Il tempo geologico è rappresentato sull’asse verticale. La tempistica degli eventi di ramificazione per i taxa è solo approssimativa e dedotta da una combinazione di dati molecolari e fossili (Steiper et al., 2004; MacLatchy et al., 2006; Steiper e Young, 2008 e 2009; Chan et al., 2010; Stevens et al., 2013). Le collocazioni filogenetiche dei taxa fossili sono ipotetiche, come discusso nel testo. La posizione di Afropithecus e Morotopithecus potrebbe essere invertita, o i due potrebbero essere taxa fratelli. NMW si riferisce alle scimmie del Nuovo Mondo e OWM si riferisce alle scimmie del Vecchio Mondo, mentre Propliopithecoidea, Pliopithecoidea e Saadanius si riferiscono ai catarrini a stelo dell’Afro-Arabia privi di sinapomorfie dei catarrini a corona.
Viste queste somiglianze adattative tra gli ominoidi viventi, è di un certo interesse indagare quali (se esistono) hanno caratterizzato la divergenza iniziale degli ominoidi da altri primati catarrini, in particolare i cercopitecoidi (scimmie del Vecchio Mondo). Le prove morfologiche e genetiche combinate collocano la divergenza ominoide-cercopitecoide a 29,6 Ma (~24-38 Ma), e la divergenza ominide-lobatide a 18,8 Ma (~16-24 Ma) (Steiper e Young, 2009). Gli ominidi dovrebbero quindi essere presenti nell’Oligocene, ma come possiamo riconoscerli? Li abbiamo trovati? L’identificazione di queste prime scimmie fossili è potenzialmente ostacolata dai seguenti problemi:
1) Un record fossile carente. Ci sono pochi siti dell’Oligocene in Africa che rappresentano il periodo appropriato, e anche se ci sono diversi siti del Miocene iniziale, molti dei loro taxa catarrini sono scarsamente rappresentati, in particolare nel postcranio.
2) La lunghezza del tempo dall’origine degli Hominoidea. I membri basali di un taxon possono avere poca somiglianza con le forme attuali perché spesso conservano numerose caratteristiche primitive. Per esempio, una scimmia molto precoce potrebbe essere stata pronogama anche se l’ultimo antenato comune delle scimmie viventi aveva sviluppato adattamenti per la postura eretta. Ci si aspetta quindi che le potenziali sinapomorfie tra gli ominoidi del tronco e della corona siano poche.
3) I lunghi periodi di evoluzione nei lignaggi esistenti possono essere stati individualmente unici, così che i taxa moderni conservano poche sinapomorfie definibili, o alcune delle loro presunte sinapomorfie possono essere parallelismi.
4) La storia evolutiva della trasformazione morfologica può non essere risolta in modo appropriato, portando a interpretazioni filogenetiche contrastanti.
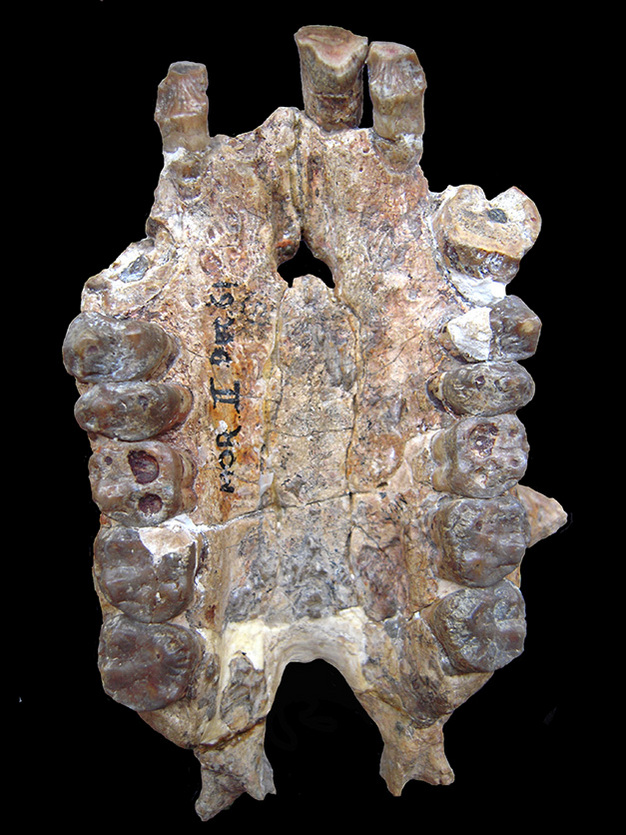
Lo studio comparativo che coinvolge sia il gambo che la corona dei catarrini può mitigare questi problemi (Zalmout et al., 2010). Per esempio, le caratteristiche presenti in una specie del tardo Oligocene o del primo Miocene, che non sono condivise primitivamente con i catarrini a stelo e i cercopitecoidi, ma sono osservate negli ominoidi a corona, sono sinapomorfie plausibili degli ominoidi. Per esempio, il seno etmo-frontale è presente nelle scimmie africane ed è assente nei catarrini a stelo afro-arabi, cercopitecoidi, Pongo e Hylobates (Rossie et al., 2002; Rossie, 2008). Il seno è presente anche in tre ominoidi putativi del primo Miocene: Proconsul, Morotopithecus e Afropithecus (ibid.). La distribuzione di questo tratto può quindi essere interpretata come un carattere derivato che si verifica dapprima negli ominoidi del tronco, e perso nelle scimmie asiatiche (Begun, 1992; Moyà-Solà e Köhler, 1995; ma vedi Rossie, 2008, per modi alternativi di interpretare questo carattere). Un’altra caratteristica cranica, una maggiore robustezza della sezione trasversale dei canini, e la concomitante espansione del palato anteriore, ha una distribuzione simile (cioè, si trova negli ominidi esistenti (persa secondariamente nell’uomo) così come in Proconsul, Morotopithecus e Afropithecus, ma è assente nei catarrini staminali e cercopitecoidi), e supporta anche l’interpretazione che i taxa del Miocene sono ominidi (Begun et al., 1997; Zalmout et al., 2010). Il Kamoyapithecus del tardo Oligocene è conosciuto solo dai resti gnatodentali (Leakey et al., 1995), ma la presenza di denti canini superiori espansi in sezione potrebbe segnalare che si tratta anche di un ominide (Zalmout et al., 2010). Inoltre, questi quattro generi fossili includono specie con dimensioni corporee stimate in eccesso di 30 kg, i primi catarrini a raggiungere queste dimensioni.
Viste le grandi dimensioni e la nicchia arborea delle scimmie esistenti (i gibboni sono probabilmente secondariamente nanizzati: Tyler, 1993), le grandi dimensioni del corpo possono anche essere una caratteristica degli ominidi del tronco. Negli ominoidi della corona, questo tratto è tradizionalmente legato allo sfruttamento da parte degli ominoidi di cibi arboricoli di alta qualità, reso possibile da innovazioni locomotorie che includono una maggiore mobilità degli arti e la capacità di afferrare (che permettono di distribuire il peso su più supporti), e l’ortogradia (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin e Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Le prove di queste trasformazioni postcraniali sono variamente espresse in Proconsul, Afropithecus e Morotopithecus. Proconsul è ben noto da diversi scheletri parziali ed è stato ricostruito come un quadrupede sopra il tronco, con un tronco pronogrado, ma sembra essere stato privo di coda (Napier e Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). Il Proconsul è anche ricostruito per avere una maggiore capacità di presa e una maggiore mobilità nelle articolazioni come il gomito, rispetto ai primitivi catarrini e cercopitecoidi, come evidenziato da caratteristiche come una zona conoidea distinta nell’omero distale (Rose, 1988). Afropithecus è meno ben rappresentato postcranialmente, ma assomiglia a Proconsul in elementi noti (Ward, 1998). Sebbene l’associazione tra i campioni cranici e postcraniali delle località di Moroto sia oggetto di disaccordo (Senut et al., 2000), i resti postcraniali di Morotopithecus differiscono in modo consistente da quelli attribuiti a Proconsul, suggerendo invece un ominoide con una parte inferiore della schiena rigida, un’articolazione gleno-omerale mobile e un femore adattato per l’arrampicata deliberata, che ricorda maggiormente gli ominoidi della corona (MacLatchy et al., 2000).
Oltre alle maggiori dimensioni del corpo, tutti i taxa che possono essere valutati hanno un pronunciato dimorfismo sessuale sia nelle dimensioni del corpo che nella morfologia canina (Kelley, 1986; Ruff et al., 1989). Mentre il forte dimorfismo sessuale è una caratteristica primitiva dei catarrini, esso è ulteriormente elaborato negli apparenti ominoidi staminali e nelle grandi scimmie moderne (ma secondariamente ridotto negli umani).
Ci sono poche, se non nessuna, sinapomorfie convincenti nella dentizione degli ominoidi. Poiché gli ominidi conservano la morfologia primitiva dei denti dei catarrini (Kay e Ungar, 1997), mentre i denti dei cercopitechi condividono la chiara sinapomorfia della bilopodontia molare, c’è una storia di utilizzo di caratteristiche dentali debolmente diagnostiche per riconoscere che alcuni catarrini dell’Oligocene e del Miocene poco rappresentati potrebbero essere ominidi, in attesa della scoperta di esemplari più completi (ad esempio Andrews, 1978 e 1992; Stevens et al., 2013). Tali catarrini oligocenici e miocenici sono stati indicati informalmente come “scimmie dentarie” (ad esempio Fleagle e Kay, 1987; Cartmill e Smith, 2009), o collocati nel limbo tassonomico utilizzando una denominazione come Superfamiglia incertae sedis (ad esempio Begun, 2007).
In particolare, è stata avanzata l’idea che nessuno dei taxa fossili discussi sopra sia un ominoide, e che il primo ominoide riconoscibile sia il Kenyapithecus di 14 milioni di anni fa, che ha alcuni attributi craniodentali derivati (ad esempio, un cingolo molare ridotto, una posizione di origine dell’arco zigomatico più alta) rispetto alle prime forme del Miocene (Harrison, 2010). Se così fosse, ci sarebbe un divario inspiegabile di almeno 10 Ma – nonostante un robusto record fossile per gran parte di questo periodo di tempo – tra l’origine stimata degli ominidi e la loro prima apparizione.
Anche se i paleoantropologi non sono in grado di determinare la posizione filogenetica delle “scimmie dentarie” o di Kamoyapithecus con fiducia, c’è un ampio sostegno all’ipotesi che Proconsul, Afropithecus e Morotopithecus siano ominoidi staminali, piuttosto che catarrini staminali. Come finestra sull’evoluzione dei primi ominoidi, questi taxa suggeriscono quanto segue:
1) Le grandi dimensioni del corpo, l’allargamento della dentizione anteriore e i sottili cambiamenti nella dentizione posteriore (ad esempio, molari più allungati e cambiamenti nella disposizione delle cuspidi molari e nello sviluppo del cingolo) suggeriscono l’occupazione di nicchie ecologiche fondamentalmente diverse da quelle dei catarrini staminali. I grandi canini funzionano sia nell’approvvigionamento di cibo che nella selezione sessuale, con un dimorfismo pronunciato che suggerisce che quest’ultima sia stata un fattore. Le grandi dimensioni permettono l’accesso a nuovi cibi e migliorano la flessibilità di foraggiamento (Wheatley, 1987), diminuiscono i rischi di predazione, promuovono la sopravvivenza e, insieme a fattori come la stabilità dell’habitat, possono essere collegati alla maturazione ritardata. L’analisi del modello di sviluppo dentale dell’Afropithecus (e forse del Proconsul) suggerisce una maturazione ritardata rispetto ad altri antropoidi (Kelley, 1997, 2002), rendendo la storia di vita lenta un’altra possibile sinapomorfia degli ominoidi.
2) All’inizio del Miocene, c’è almeno una modesta diversità locomotoria e dietetica tra i probabili ominoidi a stelo dal corpo grande, indicando l’inizio di una vera e propria radiazione adattativa, con l’aumento delle dimensioni del corpo, la crescente novità locomotoria e lo sfruttamento più specializzato delle fonti di cibo arboricole che si evolvono contemporaneamente.
3) Non è attualmente risolvibile quali caratteristiche anatomiche associate all’ortogradia e alla mobilità articolare nelle scimmie moderne siano omologhe o omoplastiche (ad esempio, Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Questo dibattito ha implicazioni per interpretare la posizione evolutiva dei presunti ominoidi staminali, e per ricostruire il modello e i tempi dell’emergere degli adattamenti delle scimmie moderne.
Per esempio, le somiglianze postcraniali tra il Morotopithecus e le scimmie viventi potrebbero essere interpretate in diversi modi. Se le scimmie viventi hanno acquisito la loro ortogonalità attraverso un’evoluzione parallela, allora il Morotopithecus rappresenta il primo caso conosciuto di una transizione che si è verificata più volte nell’evoluzione degli ominoidi. Questo è di grande interesse adattativo, suggerendo che qualche propensione genetica o di sviluppo sottostante (vedi sotto) ha continuato a guidare la risposta evolutiva degli ominoidi nella stessa direzione. Significherebbe anche che certi tratti postcraniali devono essere usati con cautela nell’analisi filogenetica (Sanders e Bodenbender, 1994). D’altra parte, se queste somiglianze locomotorie fossero ereditate da un antenato comune, il Morotopithecus sarebbe più strettamente legato agli ominoidi della corona rispetto al Proconsul (e forse all’Afropithecus). Un’altra possibilità è che Morotopithecus fosse un “esperimento” indipendente, e che l’antenato comune degli ominoidi della corona fosse comunque ortogrado.
4) Come discusso sopra, nonostante le differenze nelle specializzazioni locomotorie e i lunghi periodi di evoluzione indipendente, tutti i taxa ominoidi sopravvissuti condividono un piano corporeo simile che li differenzia dagli altri primati (con l’eccezione di alcune scimmie ateline, che hanno evoluto convergentemente torsi e specializzazioni degli arti anteriori simili a quelli delle scimmie (Erikson, 1963; Larson, 1998). Le basi anatomiche di base del piano corporeo delle scimmie hanno portato a una notevole diversità e versatilità locomotoria, consentendo ai primati dal corpo grande di muoversi efficacemente in modo arboreo (attraverso l’arrampicata e la sospensione) e terrestre (usando il knuckle-walking e il bipedalismo). Tale innovazione locomotoria, legata a caratteristiche come l’ortogradia, è probabilmente alla base del “successo” evolutivo di tutti gli ominoidi viventi, e sembra essere stata presente in qualche misura negli ominoidi staminali. Il lavoro di sviluppo di Young e colleghi (2010) ha dimostrato che le scimmie attuali hanno una ridotta integrazione tra gli arti anteriori e posteriori rispetto alle scimmie quadrupedi. Il rilassamento di questo vincolo genetico è stato interpretato per aver permesso agli arti di evolversi più liberamente in modi diversi in risposta alle pressioni di selezione (ibid.). Inferito uso differenziale degli arti in Morotopithecus (MacLatchy et al., 2013) e più giovani taxa Miocene (ad esempio Moya Sola e Kohler, 1996; Moya Sola et al, 2004; Nakatsukasa e Kunimatsu, 2009) supporta provvisoriamente la supposizione di Young e colleghi (2010) che l'”evolvibilità” degli arti possa essere un tratto ancestrale degli ominidi, ma sono necessari più materiale fossile e una migliore comprensione della genetica alla base di questo tratto.
Il futuro approfondimento delle origini degli ominidi e della prima evoluzione emergerà dai continui tentativi di identificare e comprendere la base delle possibili sinapomorfie ominidi. Questo, a sua volta, richiederà una migliore documentazione fossile sia per i catarrini a gambo che per quelli a corona, e uno studio anatomico, evolutivo e genetico più dettagliato dei caratteri condivisi in esame. Lo studio integrato degli ominoidi fossili nel loro contesto paleoambientale può anche aiutarci a capire meglio perché le caratteristiche degli ominoidi sono state selezionate, una volta o più volte.