T.H. Huxleyは他の猿を見ることは「自己のぼやけたコピー」を見ることだと書いた(1863 pg.73)。 類人猿、すなわちホミノイドの間の密接な身体的関係は、カロルス・リンネウス(1759年)により、すでに1世紀前に認識されていたのである。 肩関節の間隔が広い胴体、硬い背中、長い前肢、可動性のある四肢関節、強い把持能力、尾がないことなどである(Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996)。 ヒトは二足歩行への二次適応の結果、比較的長い腕など、これらの特徴のいくつかを欠いている。 頭蓋の特徴もホミノイドに共通しており、広い前口蓋や比較的大きな脳がある(Jerison, 1973; Rae, 1997)。 また、ホミノイドは成熟が遅れて寿命が延び(Kelley, 1997)、ヒト以外のすべての種が熱帯林に生息し、熟した果実や葉を食料としている(Fleagle, 2013)など、生態的ニッチも類似している。
ヒヨケムシ(ギボンとシャマン)は他の生きた類人猿よりも小さく、他の点でも区別されており、より最近の共通祖先を共有するヒト科(類人猿とヒト)から長い間進化的に隔離されていたことを反映しています(図1)。 ヒト科の動物は、前口蓋がヒロバット科の動物よりも広く、背が低く、硬く、上腕二頭筋の解剖学的特殊化を欠いている(Young, 2003)。
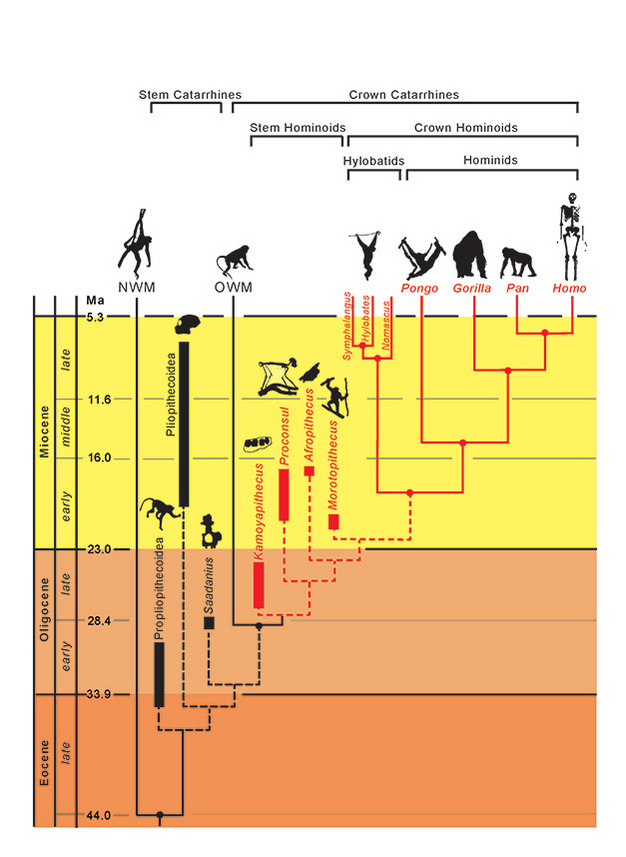
分類群の系統関係は実線(現存)および破線(絶滅)で表現されています。 縦軸は地質学的な時間を表す。 分類群の分岐時期は分子データと化石データの組み合わせから推定した近似値である(Steiper et al., 2004; MacLatchy et al., 2006; Steiper and Young, 2008 and 2009; Chan et al., 2010; Stevens et al., 2013)。 化石分類群の系統的位置づけは、本文で述べたように仮説に基づくものである。 アフロピテクスとモロトピテクスの位置は逆転する可能性もあるし、両者が姉妹分類群になる可能性もある。 NMWは新世界ザル、OWMは旧世界ザルを指し、Propliopithecoidea、Pliopithecoidea、Saadaniusは冠型カタリ派の同型を欠くアフロ・アラビア産幹型カタリ派を指す。
生きたホミノイドにこうした適応上の類似性があることから、他の双弓類、特に旧世界ザルからの最初の分岐の特徴は何か(あれば)調べることが興味深くなっています。 形態学的証拠と遺伝学的証拠を総合すると、ホミノイドとオナガザルの分岐は29.6 Ma(〜24〜38 Ma)、ヒイロバチの分岐は18.8 Ma(〜16〜24 Ma)である(Steiper and Young, 2009)。 したがって、ホミノイドは漸新世には存在しているはずであるが、どのように認識すればよいのだろうか? しかし、どのように見分ければよいのだろうか。 これらの初期類人猿の同定は、以下の問題によって妨げられる可能性がある。
1) 化石記録が不十分であること。 アフリカには適切な時期を示す漸新世の遺跡がほとんどなく、初期中新世の遺跡はいくつかあるが、そのカタロニア系分類群の多くは、特に後頭部での表現が乏しい。
2)ホミノア科の起源からの時間の長さ。 原始的な特徴を多く残しているため、現生種とあまり似ていないことがある。 例えば、類人猿の最終共通祖先が直立姿勢の適応を進化させていたとしても、ごく初期の類人猿は前かがみであった可能性がある。 従って、幹類と冠類の間で同型化される可能性は少ないと思われる。
3)現存する系統の進化の長い期間は、現代の分類群が決定的な同形態をほとんど保持しないように、個々にユニークであったか、または推定される同形態の一部が並列である可能性がある。
4)形態的変化の進化史が適切に解決されず、系統学的解釈の矛盾を招く可能性がある。
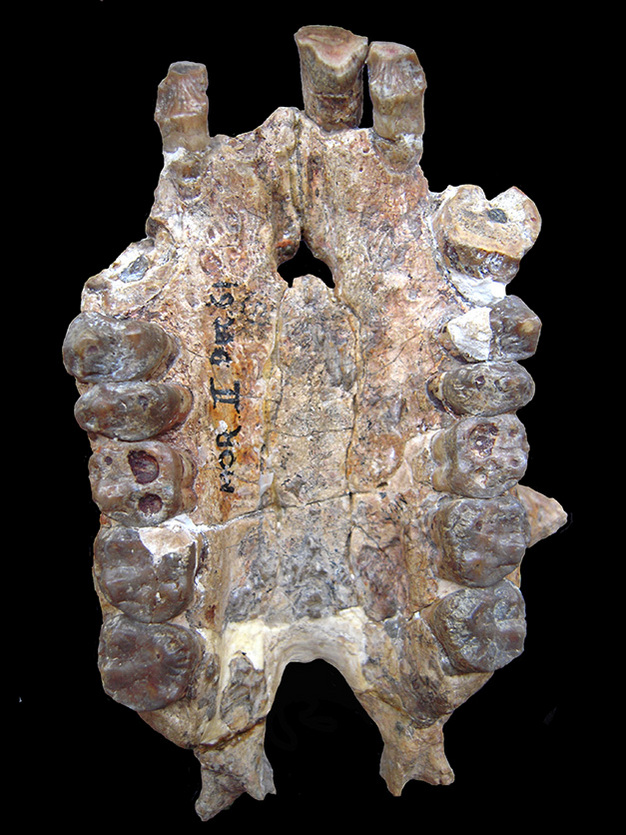
幹と冠のカタバミの両方を含む比較研究は、これらの問題を軽減できる(Zalmout他、2010)。 例えば、後期漸新統や前期中新統の種に存在する特徴で、幹カタリナ類やクマバチ類と原始的に共有されていないが、冠型ホミノイドに観察されるものは、ホミノイドの共感覚としてもっともらしいと言える。 例えば、ethmo-frontal sinusはアフリカ類人猿に存在し、アフロアラビアのstem catarrhines、cercopithecoids、Pongo、Hylobatesには存在しない(Rossie et al.2002; Rossie, 2008)。 また、洞は中新世初期の3種のホミノイドにも存在する。 Proconsul, Morotopithecus, Afropithecus (同上). したがって、この形質の分布は、幹のホミノイドに最初に生じ、アジアの類人猿で失われた派生的な性質と解釈できる(Bebun, 1992; Moyà-Solà and Köhler, 1995; ただしこの性質の別の解釈については Rossie, 2008 を参照)。 もう一つの頭蓋特徴である犬歯の断面形状の頑強化とそれに伴う前口蓋の拡大も同様の分布を示す(すなわち, 現生人類(ヒトでは二次的に失われる)や Proconsul, Morotopithecus, Afropithecus に認められ、幹のカタリナ類やcercopithecoids には認められない)、また中新世の分類群がホミニドであるという解釈(Bgun 他, 1997; Zalmout 他, 2010)も支持される。 後期漸新世のカモヤピテクスは、鼻歯遺体からしか知られていないが(Leakey et al.、1995)、断面的に拡大した上顎犬歯の存在は、この種もホミノイドであることを示唆する(Zalmout et al.) また、これら4属の化石には推定体長30kgを超える種が含まれており、カタユウレイボヤとしては初めてこのサイズに到達した。
現存する類人猿の大型化と樹上ニッチ(テナガザルは二次矮小化:Tyler, 1993)を考えると、大型化は幹ホミノイドの特徴でもあるのかもしれません。 冠位ホミノイドでは、この特徴は伝統的に、強化された四肢の運動性と把持能力(複数の支持体に体重を分散できる)、および直交性を含む運動器の革新によって可能となった、高品質の樹上食のホミノイド搾取と結びついている(Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin and Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004)。
これらの頭蓋後部の変形の証拠は、プロコンシュール、アフロピテクス、モロトピテクスで多様に表現されている。 Proconsulはいくつかの部分骨格からよく知られており、前向きの体幹を持つ前枝のある四足動物として復元されているが、尾を欠いていたようである(Napier and Davis, 1959; Beard et al.) また、上腕骨遠位部に明瞭な円錐角部(zona conoidea)があることから、原始的なカタリナ類やオナガザル類に比べ、把握能力が高く、肘などの関節に大きな可動性があったと推定されている(Rose, 1988)。 Afropithecusは頭蓋後部の表現が少ないが、知られている要素ではProconsulに似ている(Ward, 1998)。 モロト出土の頭蓋と後頭蓋の関連性については異論があるが(Senut et al, 2000)。
大きな体格に加えて、評価可能なすべての分類群には、体格と犬歯の形態の両方で顕著な性的二型がある(Kelley、1986;Ruffら、1989)。 2239>ホミノイドの歯列には、説得力のある同型異形がほとんどない。 ホミノイドは原始的なカタリニアの歯の形態を保持しており(Kay and Ungar, 1997)、一方、オナガザルの歯は臼歯両生類という明確な同型性を共有しているので、より完全な標本の発見を待って、弱く診断できる歯の特徴を使って、いくつかの漸新統や中新統のカタリニア人がホミノイドかもしれないという歴史がある(例: Andrews, 1978 and 1992; Stevens et al, 2013)。 このような漸新世・中新世のカタユウレイボヤは、非公式に「歯類人猿」と呼ばれたり(Fleagle and Kay, 1987; Cartmill and Smith, 2009)、Superfamily incertae sedis などと呼ばれて分類学的に宙ぶらりんになっている(Begun, 2007 など)。
注目すべきは、上記の化石分類群のどれもがホミノイドではなく、1400万年前のケニアピテクスが最初のホミノイドと認識され、初期中新世に比べ、いくつかの頭蓋の属性(例:臼歯部の縮小、頬骨弓の高い位置の起源)を持っているという考え方です(Harrison、2010年)。 古人類学者は、「歯の類人猿」やカモヤピテクスの系統分類を確信を持って決定することはできないが、プロコンスル、アフロピテクス、モロトピテクスは、幹のカタリ派ではなく幹のホミノ派だという仮説を広く支持している。 ホミノイドの初期進化を知る窓として、これらの分類群は次のことを示唆している。
1)大きな体格、前歯部の拡大、後歯部の微妙な変化(例えば、より細長い臼歯、臼歯の尖端配置や稜鞘の発達の変化)は、幹型ネコ科動物とは根本的に異なる生態的ニッチの占有を示唆するものであった。 大型犬歯は食料調達と性淘汰の両面で機能し、顕著な二型性は後者が要因であったことを示唆する。 大型化は新しい餌へのアクセスを可能にし、採餌の柔軟性を向上させ(Wheatley, 1987)、捕食のリスクを減らし、生存率を高め、生息地の安定性などの要因と相まって、成熟の遅延と関連している可能性がある。 アフロピテクス(とおそらくプロコンシュール)の歯列発達パターンの分析から、他の類人猿に比べて成熟が遅いことが示唆され(Kelley, 1997, 2002)、遅い生活史も幹ホミノイドの共形態の一つである可能性が出てきた。
2) 中新世初期には、大型の幹ホミノイドの間で少なくとも適度な運動と食事の多様性があり、真の適応放散の始まりを示し、身体サイズの増加、運動器の新規性の増加、樹上食料源のより専門化した利用が同時に進化している。
3) 現代類人猿の直交性と関節可動性に関連する解剖学的特徴が、相同性か相塑性かは現在のところ解決されていない(例えば、Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoyら, 2009)。 この議論は、幹とされるホミノイドの進化的位置づけの解釈や、現代の猿の適応の出現のパターンや時期の再構築に影響を与えるものです。 もし類人猿が並行進化によって直交性を獲得したのであれば、モロトピテクスはホミノイドの進化において何度か起こった変遷の最も早い例を示していることになる。 このことは、ホミノイドの進化を同じ方向に誘導し続けた、何らかの発生的・遺伝的傾向(後述)を示唆し、適応上非常に興味深いことである。 また、ある種の後頭骨形質が系統分析に慎重に使用されなければならないことも意味する(Sanders and Bodenbender, 1994)。 一方、これらの運動器の類似性が共通の祖先から受け継がれたとすれば、モロトピテクスはプロコンスル(そしておそらくアフロピテクス)よりも冠歯類に近縁であることになる。 さらに別の可能性としては、モロトピテクスは独立した「実験」であり、それにもかかわらずクラウンホミノイドの共通祖先は直交性であったということです。
4) 以上のように、運動器の特殊性の違いや独立した進化の長い期間にもかかわらず、現存するすべてのホミノイド分類群は、他の霊長類とは異なる同様の身体計画を共有している(ただし、一部のアテネザルは、猿人と同様の胴体と前肢の特殊性を収斂進化させている(Erikson、1963;Larson、1998))。 このような猿のボディプランの基本的な解剖学的基盤が、驚くべき運動多様性と多用途性をもたらし、大きな体の霊長類が、樹上移動(クライミングとサスペンションによる)と地上移動(ナックルウォークと二足歩行による)を効果的に行うことを可能にしているのである。 このような運動器の革新性は、直交性などの特徴と結びついて、現存するすべてのホミノイドの進化の「成功」の根底にあると思われ、幹ホミノイドにもある程度は存在していたようである。 Youngら(2010)による発生学的研究により、現存する類人猿は四足歩行のサルと比較して前肢と後肢の統合性が低下していることが示されている。 この遺伝的制約が緩和されたことで、四肢は淘汰圧に応じてより自由に異なる進化を遂げるようになったと解釈されている(ibid.)。 モロトピテクス (MacLatchy et al., 2013) や中新世の若い分類群では、四肢の使い分けが推定されている (例えば、Moya Sola and Kohler, 1996; Moya Sola et al…, ホミノイドの起源と初期進化に関する将来の洞察は、ホミノイドの共形態の可能性の根拠を特定し理解する試みを継続することから生まれるだろう。 そのためには、幹と冠の両方のカタリニアの化石記録を改善し、精査中の共有文字の解剖学的、発生学的、遺伝学的研究をより詳細に行うことが必要であろう。 また、ホミノイドの化石を古環境の文脈で統合的に研究することで、なぜホミノイドの特徴が一度や二度選択されたのかをより理解することができるかもしれない
。