Odhalte proces prokaryotické transkripce a translace
Prokaryota, mezi něž patří bakterie a archea, jsou většinou jednobuněčné organismy, které z definice postrádají membránová jádra a další organely. Bakteriální chromozom je kovalentně uzavřený kruh, který na rozdíl od eukaryotických chromozomů není uspořádán kolem histonových proteinů. Centrální oblast buňky, v níž se nachází prokaryotická DNA, se nazývá nukleoid. Kromě toho mají prokaryota často hojné plazmidy, což jsou kratší kruhové molekuly DNA, které mohou obsahovat pouze jeden nebo několik málo genů. Plazmidy se mohou přenášet nezávisle na bakteriálním chromozomu během buněčného dělení a často nesou vlastnosti, jako je odolnost vůči antibiotikům. Kvůli těmto jedinečným vlastnostem se transkripce a regulace genů u prokaryotických a eukaryotických buněk poněkud liší.
Cíle učení
- Pochopit základní kroky při přepisu DNA do RNA v prokaryotických buňkách
- Pochopit základy prokaryotické translace. a jak se liší od eukaryotické translace
Prokaryotická transkripce
Iniciace transkripce u prokaryot
Prokaryota nemají membránovou transkripci.uzavřená jádra. Proto mohou procesy transkripce, translace a degradace mRNA probíhat současně. Intracelulární hladina bakteriálního proteinu může být rychle zesílena více transkripčními a translačními událostmi probíhajícími současně na stejném templátu DNA. Prokaryotická transkripce často pokrývá více než jeden gen a vytváří polycistronní mRNA, která specifikuje více než jeden protein.
Naše diskuse zde bude ilustrovat transkripci popisem tohoto procesu u Escherichia coli, dobře prostudovaného bakteriálního druhu. Přestože mezi transkripcí u E. coli a transkripcí u archeí existují určité rozdíly, lze pochopení transkripce u E. coli aplikovat prakticky na všechny bakteriální druhy.
Prokaryotická RNA polymeráza
Prokaryota používají k transkripci všech svých genů stejnou RNA polymerázu. U E. coli se polymeráza skládá z pěti polypeptidových podjednotek, z nichž dvě jsou identické. Čtyři z těchto podjednotek, označované α, α, β a β′, tvoří jádro enzymu polymerázy. Tyto podjednotky se sestavují při každém přepisu genu a po dokončení přepisu se rozloží. Každá podjednotka má jedinečnou úlohu; dvě podjednotky α jsou nezbytné pro sestavení polymerázy na DNA; podjednotka β se váže na ribonukleosidtrifosfát, který se stane součástí vznikající „čerstvě zrozené“ molekuly mRNA; a β′ váže templátové vlákno DNA. Pátá podjednotka, σ, se podílí pouze na iniciaci transkripce. Propůjčuje transkripční specifitu, takže polymeráza začne syntetizovat mRNA z příslušného iniciačního místa. Bez σ by základní enzym přepisoval z náhodných míst a vytvářel by molekuly mRNA, které by specifikovaly proteinovou hatmatilku. Polymeráza složená ze všech pěti podjednotek se nazývá holoenzym (holoenzym je biochemicky aktivní sloučenina složená z enzymu a jeho koenzymu).
Prokaryotické promotory

Obrázek 1. Podjednotka σ prokaryotické RNA polymerázy rozpoznává konsensuální sekvence, které se nacházejí v promotorové oblasti před startem transkripce. Podjednotka σ se od polymerázy oddělí po zahájení transkripce.
Promotor je sekvence DNA, na kterou se naváže transkripční stroj a zahájí transkripci. Ve většině případů existují promotory před geny, které regulují. Specifická sekvence promotoru je velmi důležitá, protože určuje, zda je příslušný gen přepisován stále, částečně nebo zřídka. Ačkoli se promotory mezi prokaryotickými genomy liší, několik prvků je konzervovaných. V oblastech -10 a -35 před iniciačním místem se nacházejí dvě promotorové konsensuální sekvence neboli oblasti, které jsou podobné u všech promotorů a u různých bakteriálních druhů (obrázek 1).
Konsensuální sekvence -10, nazývaná oblast -10, je TATAAT. Sekvence -35, TTGACA, je rozpoznávána a vázána σ. Jakmile dojde k této interakci, podjednotky jádra enzymu se na toto místo navážou. Oblast -10 bohatá na A-T usnadňuje odvíjení templátu DNA a vzniká několik fosfodiesterových vazeb. Iniciační fáze transkripce končí tvorbou abortivních transkriptů, což jsou polymery o délce přibližně 10 nukleotidů, které jsou vytvořeny a uvolněny.
Prodlužování a ukončování u prokaryot
Fáze prodlužování transkripce začíná uvolněním podjednotky σ z polymerázy. Disociace σ umožňuje jádru enzymu postupovat podél templátu DNA a syntetizovat mRNA ve směru 5′ až 3′ rychlostí přibližně 40 nukleotidů za sekundu. Jak elongace probíhá, DNA se neustále odvíjí před jádrovým enzymem a navíjí se za ním (obrázek 2). Párování bází mezi DNA a RNA není dostatečně stabilní, aby udrželo stabilitu složek syntézy mRNA. Místo toho polymeráza RNA funguje jako stabilní spojovací článek mezi templátem DNA a vznikajícími vlákny RNA, aby se zajistilo, že elongace nebude předčasně přerušena.

Obrázek 2. Kliknutím získáte větší obrázek. Během elongace prokaryotická polymeráza RNA sleduje podél templátu DNA, syntetizuje mRNA ve směru od 5′ k 3′ a odvíjí a převíjí DNA podle toho, jak je čtena.
Prokaryotické terminační signály
Po přepisu genu je třeba dát prokaryotické polymeráze pokyn, aby se oddělila od templátu DNA a uvolnila nově vytvořenou mRNA. V závislosti na přepisovaném genu existují dva druhy terminačních signálů. Jeden je na bázi bílkovin a druhý na bázi RNA. Terminace závislá na proteinu rho je řízena proteinem rho, který se sleduje za polymerázou na rostoucím řetězci mRNA. Blízko konce genu narazí polymeráza na řetězec nukleotidů G na templátu DNA a zastaví se. V důsledku toho se protein rho srazí s polymerázou. Interakce s rho uvolní mRNA z transkripční bubliny.
Ukončení nezávislé na rho je řízeno specifickými sekvencemi v templátovém řetězci DNA. Když se polymeráza blíží ke konci přepisovaného genu, narazí na oblast bohatou na C-G nukleotidy. MRNA se složí zpět na sebe a komplementární C-G nukleotidy se spojí. Výsledkem je stabilní vlásenka, která způsobí, že se polymeráza zastaví, jakmile začne přepisovat oblast bohatou na A-T nukleotidy. Komplementární oblast U-A transkriptu mRNA vytváří pouze slabou interakci s templátovou DNA. To spolu se zastavenou polymerázou vyvolá dostatečnou nestabilitu, aby se jádro enzymu odtrhlo a uvolnilo nový transkript mRNA.
Po ukončení je proces transkripce dokončen. V době, kdy dojde k terminaci, by již byl prokaryotický transkript použit k zahájení syntézy mnoha kopií kódovaného proteinu, protože tyto procesy mohou probíhat současně. Sjednocení transkripce, translace, a dokonce i degradace mRNA je možné, protože všechny tyto procesy probíhají ve stejném směru 5′ až 3′ a protože v prokaryotické buňce neexistuje membránová kompartmentalizace (obrázek 3). Naproti tomu přítomnost jádra v eukaryotických buňkách vylučuje současnou transkripci a translaci.

Obrázek 3. Více polymeráz může přepisovat jeden bakteriální gen, zatímco četné ribosomy současně překládají transkripty mRNA na polypeptidy. Tímto způsobem může určitý protein rychle dosáhnout vysoké koncentrace v bakteriální buňce.
Podívejte se na tuto animaci BioStudia a prohlédněte si proces prokaryotické transkripce.
Pracovní otázky
Která z podjednotek E. coli polymerázy propůjčuje transkripci specifičnost?
- α
- β
- β′
- σ
Oblasti -10 a -35 prokaryotických promotorů se nazývají konsenzuální sekvence, protože ________.
- jsou identické u všech bakteriálních druhů
- jsou podobné u všech bakteriálních druhů
- existují u všech organismů
- mají stejnou funkci u všech organismů
Prokaryotická translace
Translace je u prokaryot a eukaryot podobná. Zde prozkoumáme, jak probíhá translace u E. coli, zástupce prokaryot, a upřesníme případné rozdíly mezi bakteriální a eukaryotickou translací.
Iniciace
Iniciace syntézy proteinů začíná vytvořením iniciačního komplexu. U E. coli tento komplex zahrnuje malý ribozom 30S, templát mRNA, tři iniciační faktory, které pomáhají správnému sestavení ribozomu, guanosintrifosfát (GTP), který funguje jako zdroj energie, a speciální iniciační tRNA nesoucí N-formylmethionin (fMet-tRNAfMet) (obr. 4). Iniciační tRNA interaguje se start kodonem AUG mRNA a nese formylovaný methionin (fMet). Kvůli své účasti na iniciaci je fMet vložen na začátek (N-konec) každého polypeptidového řetězce syntetizovaného E. coli. V mRNA E. coli je před prvním kodonem AUG vedoucí sekvence, nazývaná Shine-Dalgarnova sekvence (známá také jako ribozomální vazebné místo AGGAGG), která interaguje prostřednictvím komplementárního párování bází s molekulami rRNA tvořícími ribozom. Tato interakce ukotvuje ribozomální podjednotku 30S na správném místě na templátu mRNA. V tomto bodě se pak ribozomální podjednotka 50S naváže na iniciační komplex a vytvoří neporušený ribozom.
U eukaryot probíhá tvorba iniciačního komplexu podobně s následujícími rozdíly:
- Iniciační tRNA je jiná specializovaná tRNA nesoucí methionin, nazývaná Met-tRNAi
- Místo vazby na mRNA v sekvenci Shine-Dalgarno rozpozná eukaryotický iniciační komplex 5′ čepičku eukaryotické mRNA a poté sleduje mRNA ve směru 5′ až 3′, dokud nerozpozná startovací kodon AUG. V tomto okamžiku se podjednotka 60S naváže na komplex Met-tRNAi, mRNA a podjednotky 40S.

Obrázek 4. Uvolnění polypeptidu. Translace u bakterií začíná vytvořením iniciačního komplexu, který zahrnuje malou ribozomální podjednotku, mRNA, iniciační tRNA nesoucí N-formylmethionin a iniciační faktory. Poté se naváže podjednotka 50S a vytvoří se neporušený ribozom.
Elongace
U prokaryot a eukaryot jsou základy elongace translace stejné. U E. coli tvoří vazba ribozomální podjednotky 50S za vzniku intaktního ribozomu tři funkčně důležitá ribozomální místa: Místo A (aminoacyl) váže příchozí nabité aminoacyl tRNA. Místo P (peptidyl) váže nabité tRNA nesoucí aminokyseliny, které vytvořily peptidové vazby s rostoucím polypeptidovým řetězcem, ale ještě se neoddělily od příslušné tRNA. Místo E (exit) uvolňuje disociované tRNA, aby mohly být znovu nabity volnými aminokyselinami. V této montážní linii tRNA existuje jedna významná výjimka: Během tvorby iniciačního komplexu vstupuje bakteriální fMet-tRNAfMet nebo eukaryotická Met-tRNAi přímo do místa P, aniž by nejprve vstoupila do místa A, čímž vzniká volné místo A připravené přijmout tRNA odpovídající prvnímu kodonu po AUG.
Prodlužování probíhá pomocí jednokodonových pohybů ribozomu, z nichž každý se nazývá translokační událost. Během každé translokační události nabitá tRNA vstupuje do místa A, pak se přesune do místa P a nakonec do místa E, odkud se odstraní. Ribozomální pohyby neboli kroky jsou vyvolány konformačními změnami, které posunou ribozom o tři báze ve směru 3′. Mezi aminoskupinou aminokyseliny připojené k tRNA v místě A a karboxylovou skupinou aminokyseliny připojené k tRNA v místě P vznikají peptidové vazby. Tvorbu každé peptidové vazby katalyzuje peptidyltransferáza, ribozym na bázi RNA, který je integrován do ribozomální podjednotky 50S. Aminokyselina navázaná na tRNA v místě P je rovněž spojena s rostoucím polypeptidovým řetězcem. Jak ribozom postupuje přes mRNA, bývalá tRNA v místě P vstupuje do místa E, odděluje se od aminokyseliny a je vyloučena. Několik kroků během elongace, včetně vazby nabité aminoacyl tRNA na místo A a translokace, vyžaduje energii získanou hydrolýzou GTP, která je katalyzována specifickými elongačními faktory. Je úžasné, že translačnímu aparátu E. coli trvá přidání každé aminokyseliny pouhých 0,05 sekundy, což znamená, že protein o 200 aminokyselinách může být přeložen za pouhých 10 sekund.
Konec
K ukončení translace dochází, když se vyskytne nesmyslný kodon (UAA, UAG nebo UGA), pro který neexistuje komplementární tRNA. Při zarovnání s místem A jsou tyto nesmyslné kodony u prokaryot a eukaryot rozpoznány uvolňovacími faktory, které způsobí, že se aminokyselina v místě P oddělí od své tRNA a uvolní nově vytvořený polypeptid. Malá a velká ribozomální podjednotka se oddělí od mRNA a od sebe navzájem; téměř okamžitě se zapojí do dalšího translačního iniciačního komplexu.
Shrnem lze říci, že existuje několik klíčových rysů, které odlišují prokaryotickou genovou expresi od exprese pozorované u eukaryot. Jsou znázorněny na obrázku 5 a uvedeny v tabulce 1.
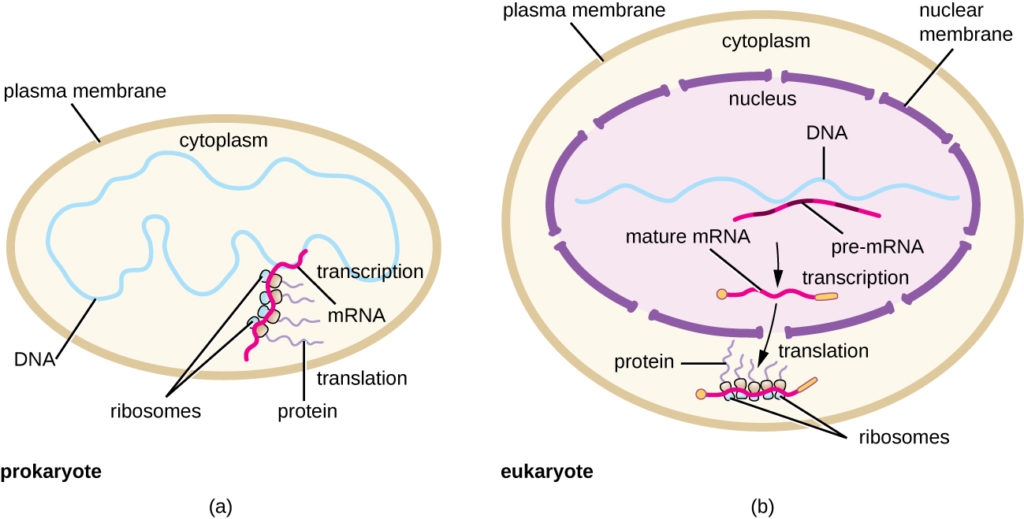
Obrázek 5. (a) U prokaryot probíhají procesy transkripce a translace v cytoplazmě současně, což umožňuje rychlou buněčnou reakci na podnět z prostředí. (b) U eukaryot je transkripce lokalizována v jádře a translace v cytoplazmě, což tyto procesy odděluje a vyžaduje zpracování RNA za účelem její stability.
| Tabulka 1. Srovnání translace u bakterií a eukaryot | ||
|---|---|---|
| Vlastnosti | Bakterie | Eukaryota |
| Ribosomy | 70S
|
80S
|
| Aminokyselina nesená iniciátorovou tRNA | fMet | Met |
| Světlo-Dalgarnova sekvence v mRNA | Přítomný | Absentní |
| Současný přepis a překlad | Ano | Ne |
Ověřte si své porozumění
Odpovězte na níže uvedené otázky, abyste zjistili, jak dobře rozumíte tématům probíraným v předchozí části. Tento krátký kvíz se nezapočítává do vašeho hodnocení v hodině a můžete jej opakovat neomezený početkrát.
Pomocí tohoto kvízu si můžete ověřit své porozumění a rozhodnout se, zda (1) budete předchozí oddíl dále studovat, nebo (2) přejdete k dalšímu oddílu.
.