Obecný úvod
Většina z nás ví, že kvasinky jsou velmi užitečný organismus, zejména pokud jde o pečení, výrobu vína a pivovarnictví. Co jsou však kvasinky a proč se na ně zaměřuje tolik výzkumu?“

Kvasinky jsou houby
Kvasinky jsou jednobuněčné mikroorganismy, které se spolu s plísněmi a houbami řadí do říše hub. Kvasinky jsou evolučně rozmanité, a proto jsou řazeny do dvou samostatných fylů, Ascomycota neboli váčkovité houby a Basidiomycota neboli vyšší houby, které společně tvoří podříši Dikarya. Pupenaté kvasinky, označované také jako „pravé kvasinky“, patří do fylogeneze Ascomycota a řádu Saccharomycetales. Tato klasifikace je založena na vlastnostech buňky, askospor a kolonie a také na buněčné fyziologii.
Kvasinky jsou jednobuněčné, ale s buněčnou organizací podobnou vyšším organismům
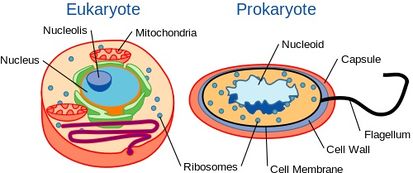
Ačkoli jsou kvasinky jednobuněčné organismy, mají podobné buněčné uspořádání jako vyšší organismy včetně člověka. Konkrétně je jejich genetický obsah obsažen v jádře. To je řadí mezi eukaryotické organismy, na rozdíl od jejich jednobuněčných protějšků, bakterií, které jádro nemají a jsou považovány za prokaryota.
Přírodní stanoviště
Kvasinky jsou v přírodě široce rozptýleny a mají širokou škálu stanovišť. Běžně se vyskytují na listech, květech a plodech rostlin a také v půdě. Kvasinky se vyskytují také na povrchu kůže a ve střevním traktu teplokrevných živočichů, kde mohou žít symbioticky nebo jako parazité. Běžnou „kvasinkovou infekci“ obvykle způsobuje Candida albicans. Kromě toho, že je Candida původcem vaginálních kvasinkových infekcí, je také příčinou opruzenin a afty v ústech a krku.
Proč studovat kvasinky?
Představte si organismus, který rychle roste v baňce a s jehož DNA lze snadno manipulovat, ale také umožňuje nahlédnout do základních lidských biologických procesů, včetně nemocí. Kvasinky tomuto popisu odpovídají a jsou předmětem studia vědců po celém světě, což vyústilo ve více než 50 000 publikovaných vědeckých článků popisujících výzkum kvasinek!

Jaké specifické vlastnosti kvasinek z nich činí „modelový organismus“ pro studium a předmět tolika výzkumů? Kvasinky jsou jednobuněčné (jednobuněčné) organismy, díky čemuž je jejich studium jednoduché, ale mají podobné buněčné uspořádání jako vyšší, mnohobuněčné organismy, například člověk – to znamená, že mají jádro, a jsou tedy eukaryoty, jak bylo popsáno výše. Nejdůležitější je, že podobnost buněčného uspořádání mezi kvasinkami a vyššími eukaryoty se promítá do podobnosti jejich základních buněčných procesů, takže objevy u kvasinek často poskytují přímé nebo nepřímé vodítko k tomu, jak biologické procesy fungují u lidí.

Další důležitou vlastností kvasinek podstatnou pro jejich roli „modelových organismů“ je skutečnost, že se s nimi relativně snadno pracuje. Kvasinky se rychle replikují a lze s nimi snadno geneticky manipulovat. Doba zdvojení kvasinek (doba potřebná k tomu, aby se buňka sama zdvojila a rozdělila) je přibližně 90 minut. Naproti tomu lidské buňky rostoucí v kultuře potřebují ke zdvojení asi 24 hodin. Pro kvasinky také existují dobře definované genetické metody, které výzkumníkům umožňují snadno izolovat mutanty, křížit je s jinými mutanty nebo na jiném genetickém pozadí a mapovat umístění genů. Genetické mapy zkonstruované na základě genetické vzdálenosti mezi geny vlastně poskytly vědcům první pohled na genom a jeho uspořádání a byly vyvrcholením genetických studií, které se datují od první poloviny dvacátého století.
Zrychlené tempo objevů bylo umožněno poté, co se genom pekařských kvasinek (S. cerevisiae), představující kompletní soubor jejich genetického materiálu, stal prvním eukaryotickým genomem, který byl sekvenován již v roce 1996. Je menší a kompaktnější než lidský genom (12 milionů párů bází a ~6 000 genů oproti 3 miliardám párů bází a ~20-25 000 genů kódujících bílkoviny). Přesto porovnání genomů ukazuje, že ~31 % kvasinkových genů je velmi podobných lidským genům a 20 % lidských chorobných genů má protějšky v kvasinkách. Kromě toho mohou kvasinkové buňky existovat buď jako haploidi (jedna sada chromozomů), nebo diploidi (dvě sady chromozomů). Protože haploidi mají pouze jednu kopii každého genu a účinné přerušení a opětovné spojení vláken DNA (rekombinace), je velmi snadné odstranit určitý gen v haploidu a pozorovat účinky na buňku neboli „fenotyp“ delečního mutanta. Diploidní buňky naproti tomu umožňují studovat esenciální geny (ty, které jsou nezbytné pro růst a životaschopnost) tím, že se jedna kopie genu odstraní a v druhé kopii se provedou jemné změny. A konečně, díky informacím ze sekvence genomu byla vytvořena rozsáhlá sada molekulárních činidel a kolekcí genomů, které vědcům poskytují výkonné prostředky ke studiu biologických problémů. Pokud je známo, že kvasinkový gen má podobnou sekvenci DNA jako lidský gen, mohou studie na kvasinkách poskytnout účinná vodítka, pokud jde o úlohu příbuzného genu (příbuzných genů) u člověka. Relativní jednoduchost studia buněčných funkcí u kvasinek v kombinaci s jejich významem pro vyšší organismy z nich tedy činí velmi účinný „modelový organismus“ pro studium.
Životní cyklus kvasinek a buněčné cykly
Kvasinky obvykle rostou nepohlavně pučením. Na mateřské (mateřské) buňce se vytvoří malý pupen, který se stane dceřinou buňkou, a s pokračujícím růstem se zvětšuje. Jak dceřiná buňka roste, mateřská buňka se duplikuje a poté segreguje svou DNA. Jádro se rozdělí a migruje do dceřiné buňky. Jakmile pupen obsahuje jádro a dosáhne určité velikosti, oddělí se od mateřské buňky. Řada událostí, které probíhají v buňce a vedou ke zdvojení a rozdělení, se označuje jako buněčný cyklus. Buněčný cyklus se skládá ze čtyř různých fází (G1, S, G2 a M) a je regulován podobně jako buněčný cyklus u větších eukaryot. Dokud je k dispozici dostatek živin, jako je cukr, dusík a fosfát, kvasinkové buňky se dělí nepohlavně.
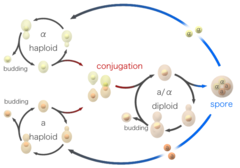
Kvasinkové buňky se mohou rozmnožovat i pohlavně. Kvasinkové buňky existují jako jeden ze dvou různých typů páření, a buňky a alfa buňky. Když se buňky opačných pářících se typů smíchají v laboratoři nebo se náhodně dostanou do kontaktu v přírodě, mohou se pářit (konjugovat). Před spojením buňky změní tvar v procesu zvaném shmooing. Termín „shmoo“ vznikl na základě tvarové podobnosti se stejnojmennou fiktivní kreslenou postavičkou, kterou koncem 40. let vytvořil Al Capp a která se poprvé objevila v jeho komiksu L’il Abner. Během konjugace se haploidní buňky shmoo nejprve spojí a poté jejich jádra splynou, čímž vznikne diploidní buňka se dvěma kopiemi každého chromozomu. Po vzniku se diploidní buňky mohou rozmnožovat nepohlavně pučením, podobně jako haploidi. Pokud však diploidní buňky postrádají živiny, dochází k jejich sporulaci. Během sporulace procházejí diploidní buňky meiózou, zvláštní formou buněčného dělení, při níž se počet chromozomů sníží ze dvou kopií zpět na jednu. Po meióze se haploidní jádra vzniklá při meióze zabalí do čtyř spor, které obsahují modifikované buněčné stěny, což vede ke vzniku struktur, které jsou velmi odolné vůči stresu prostředí. Tyto spory mohou přežívat dlouhou dobu, dokud nenastanou příznivější podmínky, například za přítomnosti lepších živin, načež jsou schopny vyklíčit a rozmnožovat se nepohlavně. Tyto různé stavy, pučení, konjugace a sporulace společně tvoří životní cyklus kvasinek.

Růst a metabolismus kvasinek
Když jsou kvasinkové buňky pěstovány v bohatých zdrojích uhlíku, jako je glukóza, preferují růst kvašením. Při kvašení se glukóza přeměňuje na oxid uhličitý a ethanol. Obecně probíhá kvašení za nepřítomnosti kyslíku, a je tedy svou povahou anaerobní. I v přítomnosti kyslíku dávají kvasinkové buňky přednost fermentačnímu růstu, což se označuje jako Crabtreeho efekt podle biologa, který tuto preferenci objevil. Tato forma růstu se využívá při výrobě chleba, piva, vína a dalších alkoholických nápojů. Ačkoli buňky kvasinek dávají přednost kvašení, při nedostatku živin jsou schopny růst i buněčným dýcháním. Při dýchání buňky přeměňují glukózu na oxid uhličitý a vodu, spotřebovávají přitom kyslík a výsledkem je produkce mnohem většího množství energie ve formě ATP.
Historické objevy

Kvasinky se jako průmyslový mikroorganismus používají již 1000 let. Staří Egypťané používali kvasinky ke kvašení chleba. Existují důkazy o mlecích kamenech, pekařských komorách a kresby 4000 let starých pekáren. Archeologické vykopávky odhalily důkazy v podobě nádob se zbytky vína starého 7 000 let.
Kvasnice poprvé zviditelnil v roce 1680 Antoni van Leeuwenhoek pomocí kvalitních čoček. Domníval se však, že tyto kuličky jsou spíše škrobovými částicemi obilí používanými k výrobě mladiny, tekutého extraktu používaného při vaření piva, než kvasnými buňkami. V roce 1789 přispěl francouzský chemik Antoine Lavoisier k pochopení základních chemických reakcí potřebných k výrobě alkoholu z cukrové třtiny. Odhadnutím podílu výchozích látek a produktů (ethanolu a oxidu uhličitého) po přidání kvasnicové pasty dospěl k závěru, že se používají dvě chemické cesty, přičemž dvě třetiny cukru se redukují na alkohol a jedna třetina na oxid uhličitý. V té době se však mělo za to, že kvasinky slouží spíše jen k zahájení reakce než k jejímu průběhu.
Francouzský chemik Joseph-Louis Gay-Lussac v roce 1815 vyvinul metody udržování hroznové šťávy v nezkvašeném stavu a zjistil, že k přeměně nezkvašené mladiny je nutné zavedení „kvasu“ (který obsahuje kvasinky), čímž prokázal význam kvasinek pro alkoholové kvašení. V roce 1835 Charles Cagniard de la Tour pomocí výkonnějšího mikroskopu prokázal, že kvasinky jsou jednobuněčné a množí se pučením. V 50. letech 19. století Louis Pasteur zjistil, že kvašené nápoje vznikají přeměnou glukózy na etanol pomocí kvasinek, a definoval kvašení jako „dýchání bez přístupu vzduchu“. Ke konci 19. století použil Eduard Buchner bezbuněčné extrakty získané rozemletím kvasinkových buněk k detekci zymázy, souboru enzymů, které podporují nebo katalyzují kvašení, za což mu byla v roce 1907 udělena Nobelova cena.
Velkou část průkopnických prací v oblasti genetiky kvasinek provedl Øjvind Winge. Objevil, že kvasinky střídají haploidní a diploidní stavy a že kvasinky jsou heterotální, protože k přeměně haploidů na diploidy (konjugaci) jsou zapotřebí dva kmeny. Spolu se svým kolegou Otto Laustsenem vymyslel techniky mikromanipulace kvasinek, aby je bylo možné geneticky zkoumat. Při této technice, známé jako „tetradová analýza“, se pomocí tenké jehly a mikroskopu izoluje struktura známá jako askus, která obsahuje čtyři spory neboli tetrády vzniklé sporulací diploida. Po izolaci askusu se spory v tetrádě oddělí a nechají se vyrůst do kolonií pro genetickou analýzu. Tato průkopnická práce mu vynesla titul „otec genetiky kvasinek“. Některé z těchto prací dále objasnil Carl Lindegren, který objasnil systém krycích typů u poupat kvasinek, prokázal existenci buněk Mat a a Mat alfa, navrhl metody, jak provádět hromadná páření mezi buňkami těchto krycích typů, a využil tyto poznatky ke studiu genetiky využití cukrů.
Od té doby provedlo mnoho dalších vědců převratný výzkum pomocí poupat kvasinek. Někteří z těchto výzkumníků byli za významné objevy učiněné během těchto studií oceněni Nobelovou cenou, mj: Dr. Leland Hartwell (2001) za objev genů, které regulují buněčný cyklus (laureát spolu s Paulem Nursem a Timem Huntem); Roger Kornberg (2006) za studie prvního kroku genové exprese, tedy způsobu, jakým se sekvence DNA genu kopíruje do messengerové RNA (mRNA); dr. Elizabeth Blackburnové, Carol Greiderové a Jacku Szostakovi (2009) za objev a objasnění genů a prostředků, kterými buňky chrání konce chromozomů neboli telomer před degradací; a doktorům Randymu Schekmanovi, Jamesi Rothmanovi a Thomasi Südhofovi (2013) za výzkum mechanismů, které regulují vezikulární provoz. Nejnověji byla cena udělena Dr. Yoshinori Ohsumimu za jeho práci o autofagii, která začala studiem kvasinek.
Komerční využití

Kvasinky jsou již dlouho považovány za organismus vhodný k výrobě alkoholických nápojů, chleba a mnoha různých průmyslových výrobků. To je založeno na snadné manipulaci s metabolismem kvasinek pomocí genetických technik, na rychlosti, s jakou je lze vypěstovat do vysokých výtěžků buněk (biomasy), na snadnosti, s jakou lze tuto biomasu oddělit od produktů, a na vědomí, že jsou obecně uznávány jako bezpečné (GRAS).
Kvasinky S. cerevisiae a další druhy kvasinek se již dlouho používají ke kvašení cukrů z rýže, pšenice, ječmene a kukuřice za účelem výroby alkoholických nápojů, jako je pivo a víno. Existují dva hlavní typy pivovarských kvasinek, kvasinky svrchního kvašení piva a kvasinky spodního kvašení ležáku. Kvasinky svrchního kvašení, jako je S. cerevisiae vystupují na povrch během kvašení a používají se k vaření piv typu ale, porter, stout a pšeničných piv. Naproti tomu S. pastorianus (dříve známá jako S. carlsbergensis) je kvasinka spodního kvašení, která se používá k výrobě ležáckého piva. Ležákové kvasinky rostou nejlépe při nižších teplotách. V důsledku toho rostou pomaleji, vytvářejí méně povrchové pěny, a proto se obvykle usazují na dně fermentoru. Plzeňské pivo, Märzen, Bocks a americké sladové likéry jsou všechny styly ležáckého piva. V moderním pivovarnictví bylo mnoho původních kmenů svrchního kvašení upraveno na kmeny spodního kvašení.
Kvasinky vyrábějí víno kvašením cukrů z hroznové šťávy (moštu) na etanol. Ačkoli kvašení vína mohou iniciovat přirozeně se vyskytující kvasinky přítomné na vinicích, mnoho vinařství se rozhodlo přidat čistou kulturu kvasinek, aby kvašení dominovaly a byly pod kontrolou. Bublinky v šampaňském a šumivých vínech vznikají sekundárním kvašením, obvykle v láhvi, které zachycuje oxid uhličitý. Oxid uhličitý vznikající při výrobě vína se uvolňuje jako vedlejší produkt. Jedna buňka kvasinek dokáže za hodinu zkvasit přibližně svou vlastní hmotnost v glukóze. Za optimálních podmínek může S. cerevisiae produkovat až 18 % objemových etanolu, přičemž normou je 15-16 %. Oxid siřičitý přítomný v komerčně vyráběném víně se přidává těsně po rozdrcení hroznů, aby zničil přirozeně přítomné bakterie, plísně a kvasinky.

Saccharomyces cerevisiae neboli pekařské droždí se již dlouho používá jako kypřící prostředek při pečení. Pekařské kvasnice fermentují cukry přítomné v těstě a produkují oxid uhličitý a etanol. Oxid uhličitý se zachytí v malých bublinkách v těstě, což způsobí, že těsto vykyne. Výjimkou je kváskový chléb, při jehož výrobě se nepoužívají pekařské kvasnice, ale kombinace divokých kvasinek a bakterií. Ke zpevnění lepku se používají kvasinky Candida milleri a ke zkvašení maltózy bakterie Lactobacillus sanfranciscensis, která vytváří kyseliny.
Kromě těchto tradičních použití se droždí používá i pro mnoho dalších komerčních aplikací. Vegani často používají droždí jako náhražku sýra a často se používá jako přísada do výrobků, jako je například popcorn. Využívá se v petrochemickém průmyslu, kde bylo upraveno pro výrobu biopaliv, jako je etanol, a farnesenu, prekurzoru nafty a leteckého paliva. Používá se také při výrobě maziv a detergentů. Kvasinky se používají v potravinářském průmyslu k výrobě potravinářských přísad včetně barviv, antioxidantů a zvýrazňovačů chuti. Často se používá při výrobě léčiv včetně antiparazitik, protinádorových látek, biofarmak, jako je inzulín, vakcín a nutraceutik. Kvasinky se běžně používají při výrobě průmyslových enzymů a chemikálií. V oblasti bioremediace životního prostředí byly kmeny dokonce využívány k odstraňování kovů z důlního odpadu.
Použití v oblasti lidských onemocnění a výzkumu

Díky vysokému stupni podobnosti kvasinkových genů s jejich lidskými protějšky a konzervované základní buněčné biologii se kvasinky staly oblíbeným modelovým systémem pro studium genů lidských onemocnění. K získání dalších informací o lidských genech bylo použito několik přístupů, jakmile bylo vytvořeno spojení mezi lidským a kvasinkovým genem. V jednom z přístupů se po objevení genu spojeného s lidským onemocněním porovnává jeho sekvence se sekvencemi všech genů v kvasinkovém genomu s cílem identifikovat nejpodobnější kvasinkový gen (geny). Pro studium, zda jsou tyto geny funkčně příbuzné, je pak lidský gen exprimován v kvasinkové skvrně, kde byl kvasinkový gen nejprve inaktivován mutací. To umožňuje výzkumníkům zjistit, zda je lidský gen schopen zachránit životaschopnost, růst nebo specifičtější defekty spojené se ztrátou kvasinkového genu, což je metoda označovaná jako funkční komplementace. Pokud jsou dráhy a/nebo procesy, kterých se kvasinkový gen účastní, zachovány, lze se o funkci lidského genu dozvědět mnohé na základě toho, co je již známo o příbuzném kvasinkovém genu. Po zavedení funkční komplementace mohou výzkumníci tento systém použít k další charakterizaci funkce produktu příbuzného lidského genu. Méně cílené přístupy, které často využívají vysoce výkonné techniky (HTP) k náhodnému screeningu tisíců lidských genů najednou s cílem identifikovat gen nebo geny s komplementární aktivitou. Takové přístupy byly úspěšně použity k identifikaci konzervovaných regulátorů buněčného cyklu (CDC2), genů zapojených do rakoviny a genů zapojených do neurodegenerativních onemocnění.
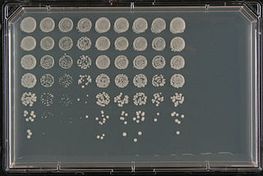
Existuje mnoho scénářů, kdy studie mohou výzkumníkům poskytnout cenné informace o buněčných drahách a/nebo procesech, do kterých je zapojen lidský gen, pokud není příbuzný kvasinkový gen přítomen. Například u některých neurodegenerativních onemocnění, jako je Alzheimerova a Parkinsonova choroba, dochází v důsledku nesprávného skládání bílkovin k hromadění proteinových agregátů zvaných amyloid, které jsou pro neurony toxické. Studium chybně složených kvasinkových proteinů s podobným potenciálem tvorby amyloidu, tzv. prionů, umožnilo vědcům nahlédnout do těchto neurodegenerativních onemocnění. Případně může zvýšená exprese genu spojeného s onemocněním v kvasinkách vést k určitému fenotypu. Například alfa-synuklein, gen spojený s Parkinsonovou chorobou, je při dostatečně vysoké expresi toxický. Takový kmen pak může být použit ke screeningu kvasinkových genů nebo malých molekul, které potlačují nebo zvyšují toxicitu vyvolanou synukleinem, což často poskytuje vodítka o příslušných buněčných drahách. Pacienti s amyotrofickou laterální sklerózou (ALS) nebo Lou Gehrigovou chorobou mají často mutace v několika proteinech vázajících RNA, což způsobuje jejich náchylnost k tvorbě agregátů, které narušují metabolismus RNA. Pomocí kvasinkového screeningu byla úspěšně identifikována řada kvasinkových genů s podobnými vlastnostmi (tvoří toxické agregáty), což vědcům poskytlo nové kandidátní geny ke studiu. Naopak lidské proteiny vázající RNA tvoří při expresi v kvasinkách toxické agregáty a tento kmen byl použit k identifikaci kvasinkového genu, který po mutaci blokuje tvorbu těchto agregátů.
Kvasinky se stávají organismem volby při studiích zaměřených na identifikaci cílů léčiv a způsobu účinku různých léčiv. Chemogenomika nebo chemická genomika označuje screeny, které využívají kombinaci chemických látek a genomiky ke zkoumání cílů léčiv a potenciální identifikaci nových léčiv. V těchto chemicko-genomických studiích se používají dva hlavní přístupy. V prvním případě je vytvořena kolekce diploidních kmenů s celým genomem, kde je jedna ze dvou identických kopií genu odstraněna, čímž se sníží hladiny určitého genového produktu. Cílové geny a geny zapojené do cílové dráhy se stávají citlivějšími na sloučeninu a jsou v tomto druhu screeningu přednostně identifikovány. Při druhém přístupu jsou systematicky odstraňovány nedůležité geny a soubor je prověřován pomocí léčiva s cílem vyhledat geny, které bufferují cílovou dráhu léčiva. Očekává se, že tento přístup umožní identifikovat geny potřebné pro růst v přítomnosti sloučeniny. Další přístupy využívající screeny s nadměrnou expresí byly použity k identifikaci genů podílejících se na rezistenci k léčivům včetně potenciálního cíle léčiva. Porovnání expresního profilu kvasinkových buněk s odstraněným genem s expresním profilem divokého typu kvasinkových buněk léčených určitým léčivem může být také účinným způsobem identifikace genů, které mohou výzkumníkům říci něco o tom, jak léčivo v buňkách funguje.
Toto je jen několik příkladů, jak lze kvasinky využít i pomoci při studiu lidských onemocnění. Studie na kvasinkách mohou výzkumníkům pomoci dozvědět se více o základní biologii pomocí tohoto modelového systému nebo jim pomoci identifikovat cíle léčiv či způsob účinku léčiv.
Zdroje
- Saccharomyces Genome Database (SGD) poskytuje komplexní integrované biologické informace pro poupata kvasinek Saccharomyces cerevisiae spolu s vyhledávacími a analytickými nástroji pro zkoumání těchto dat.
- MIPS Comprehensive Yeast Genome Database (CYPD) představuje informace o molekulární struktuře a funkční síti kompletně sekvenovaného, dobře prozkoumaného modelového eukaryota, poupátkové kvasinky Saccharomyces cerevisiae.
- Candida Genome Database (CGD), zdroj dat o genomové sekvenci a informací o genech a proteinech pro Candida albicans.
- PomBase, komplexní databáze pro štěpnou kvasinku Schizosaccharomyces pombe, která poskytuje strukturní a funkční anotace, kurátorství literatury a přístup k rozsáhlým souborům dat.
- Fungal genome resources at NCBI, průvodce zdroji houbových genomů v Národním centru pro biotechnologické informace (NCBI).
- Rozsáhlý seznam zdrojů týkajících se kvasinek na různá témata od obecných informací o kvasinkách přes nukleové kyseliny, genomy a proteiny, expresní data, lokalizaci, fenotypy a další.
Navržená četba
Knihy
- YeastBook. (2011) Obsáhlý sborník recenzí, který představuje současný stav poznání molekulární biologie, buněčné biologie a genetiky kvasinek Saccharomyces cerevisiae, Genetika
- Od a do alfa: Kvasinky jako model buněčné diferenciace. (2007) Hiten D. Madhani, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Landmark Papers in Yeast Biology. (2006), edited by Patrick Linder, David Shore, and Michael N. Hall, Cold Spring Harbor Press, Cold Spring Harbor, New York.
- Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual. (2005) David C. Amberg, Daniel J. Burke a Jeffrey N. Strathern. Cold Spring Laboratory Press, Cold Spring Harbor, New York.
- Počátky genetiky kvasinek. (1993) editoři Michael N. Hall a Patrick Linder. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Volume I: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: I.: Genome Dynamics, Protein Synthesis, and Energetics (Dynamika genomu, syntéza proteinů a energetika). (1991) edited by James R. Broach, John R. Pringle, and Elizabeth W. Jones. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Volume II: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Exprese genů. (1992) edited by Elizabeth W. Jones, John R. Pringle and James R. Broach. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Volume III: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Cell Cycle and Cell Biology (Buněčný cyklus a buněčná biologie). (1997) edited by John R. Pringle, James R. Broach and Elizabeth W. Jones. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Kvasinky: A Practical Approach. (1988) edited by I. Campbell and , and John H. Duffus, IRL Press, Ithaca, New York.
Články v časopisech
- Duina A.A., Miller M.E., and J.B. Keeney (2014) Budding Yeast for Budding Geneticists: A Primer on the Saccharomyces cerevisiae Model System. Genetika. 197:33-48.
- Botstein D. a G.R. Fink (2011) Yeast: An Experimental Organism for 21st Century Biology (Experimentální organismus pro biologii 21. století). Genetics. 189:695-704.
- Cherry J.M., Hong E.L., Amundsen C., Balakrishnan R., Binkley G., Chan E.T., Christie K.R., Costanzo M.C., Dwight S.S., Engel S.R., Fisk D.G., Hirschman J.E., Hitz B.C., Karra K., Krieger C.J., Miyasato S.R., Nash R.S., Park J., Skrzypek M.S., Simison M., Weng S. a E.D. Wong (2011) Saccharomyces Genome Database: the genomics resource of budding yeast. Nucleic Acids Res. 40:D700-D705.
- Altman T.J., Boone C., Churchill G.A., Hengartner M.O., Mackay T.F., and D.L. Stemple (2011) The future of model organisms in human disease research. Nat. Rev. Genet. 18:575-582.
- Ho C.H., Piotrowski J., Dixon S.J., Baryshnikova A., Costanzo M., and C. Boone (2011) Combining functional genomics and chemical biology to identify targets of bioactive compounds. Curr Opin Chem Biol. 15:66-78.
- Smith A.M., Ammar R., Nislow C., and G. Giaever (2010) A survey of yeast genomic assays for drug and target discovery. Pharmacol Ther. 127:156-164.
- Dolinski K., and D. Botstein (2007) Orthology and functional conservation in eukaryotes. Annu. Rev. Genet. 41:465-507.
- Barnett J.A. (2007) A history of research on yeasts 10: foundations of yeast genetics. Yeast 24:799-845.
- Spradling A., Ganetsky B., Hieter P., Johnston M., Olson M., Orr-Weaver T., Rossant J., Sanchez A. a R. Waterston (2006) New roles for model genetic organisms in understanding and treating human disease: report from the 2006 Genetics Society of America meeting. Genetics 172:2025-2032.
- Sherman F. (2002) Getting started with yeast Methods Enzymol. 350:3-41.