T.H. Huxley schrieb, dass man beim Anblick anderer Menschenaffen „verschwommene Kopien seiner selbst“ sieht (1863, S. 73). Die enge körperliche Verwandtschaft der Menschenaffen, oder Hominoiden, wurde bereits ein Jahrhundert zuvor von Carolus Linnaeus (1759) erkannt. Die lebenden Hominoiden sind durch Merkmale vereint, die mit der gewohnheitsmäßigen Orthogradie und dem Verhalten unter der Gliedmaße zusammenhängen: breiter Rumpf mit weit auseinander liegenden Schultergelenken, steifer Rücken, lange Vordergliedmaßen, bewegliche Gliedmaßen, starke Greiffähigkeit und das Fehlen eines Schwanzes (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Dem Menschen fehlen mehrere dieser Merkmale, wie z. B. relativ lange Arme, was auf die sekundäre Anpassung an den Bipedalismus zurückzuführen ist. Auch die Schädelmerkmale vereinen die Hominoiden, darunter breite vordere Gaumen und relativ große Gehirne (Jerison, 1973; Rae, 1997). Hominoide haben auch eine längere Lebensspanne mit verzögerter Reifung (Kelley, 1997) und besetzen ähnliche ökologische Nischen – alle Arten außer dem Menschen bewohnen tropische Wälder und sind in hohem Maße auf reife Früchte und Blätter als Nahrung angewiesen (Fleagle, 2013).
Hylobatiden (Gibbons und Siamangs) sind kleiner als andere lebende Menschenaffen und unterscheiden sich auch in anderer Hinsicht, was ihre lange evolutionäre Isolation von Hominiden (Menschenaffen und Menschen) widerspiegelt, die einen jüngeren gemeinsamen Vorfahren haben (Abbildung 1). Hominiden (mit Ausnahme des Menschen) haben vordere Gaumen, die breiter sind als die der Hylobatiden (Rae, 1997), besitzen noch kürzere, steifere untere Rückenpartien und haben keine anatomischen Spezialisierungen für die ricochetale Brachiation (Young, 2003).
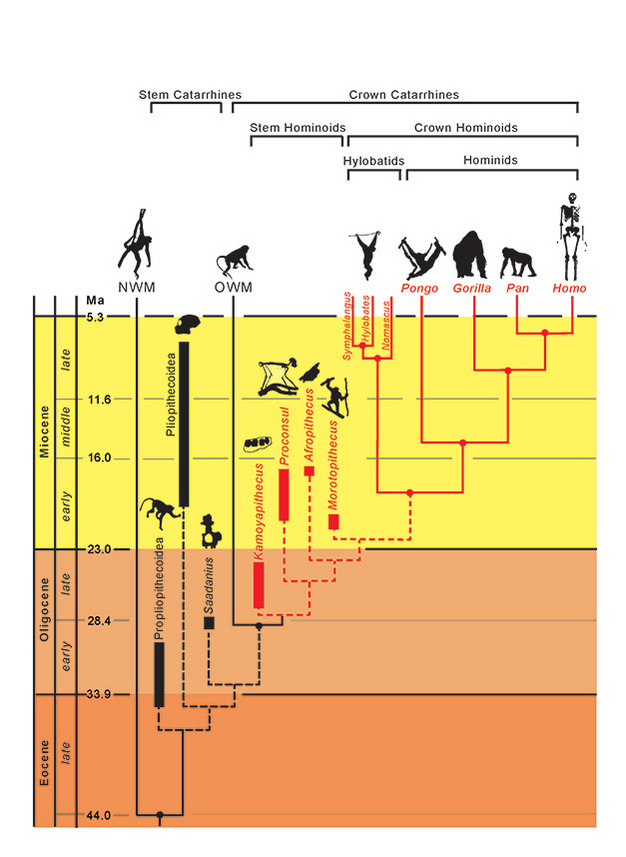
Die phylogenetischen Beziehungen der Taxa werden durch durchgezogene (ausgestorbene) und gestrichelte (ausgestorbene) Linien dargestellt. Die geologische Zeit wird auf der vertikalen Achse dargestellt. Der Zeitpunkt der Verzweigungsereignisse für Taxa ist nur annähernd und wurde aus einer Kombination von molekularen und fossilen Daten abgeleitet (Steiper et al., 2004; MacLatchy et al., 2006; Steiper und Young, 2008 und 2009; Chan et al., 2010; Stevens et al., 2013). Die phylogenetische Einordnung der fossilen Taxa ist hypothetisch, wie im Text erläutert. Die Position von Afropithecus und Morotopithecus könnte vertauscht sein, oder die beiden könnten Schwestertaxa sein. NMW bezieht sich auf Neuweltaffen und OWM auf Altweltaffen, während Propliopithecoidea, Pliopithecoidea und Saadanius sich auf Stammkatarrhinen aus Afro-Arabien beziehen, denen jegliche Kronenkatarrhinen-Synapomorphien fehlen.
Angesichts dieser adaptiven Ähnlichkeiten unter den lebenden Hominoiden ist es von einigem Interesse zu untersuchen, welche (wenn überhaupt) die anfängliche Divergenz der Hominoiden von anderen katarrhinen Primaten, insbesondere den Cercopithecoiden (Altweltaffen), kennzeichnete. Kombinierte morphologische und genetische Nachweise belegen die Divergenz zwischen Hominoiden und Cercopithecoiden bei 29,6 Ma (~24-38 Ma) und die Divergenz zwischen Hylobatiden und Hominoiden bei 18,8 Ma (~16-24 Ma) (Steiper und Young, 2009). Hominoide sollten also bereits im Oligozän vorhanden sein, aber wie können wir sie erkennen? Haben wir sie gefunden? Die Identifizierung dieser frühen fossilen Affen wird möglicherweise durch die folgenden Probleme behindert:
1) Ein unzureichender Fossilnachweis. Es gibt nur wenige oligozäne Fundstellen in Afrika, die den entsprechenden Zeitraum repräsentieren, und obwohl es mehrere Fundstellen aus dem frühen Miozän gibt, sind viele ihrer katarrhinen Taxa nur schwach vertreten, insbesondere im Postkranium.
2) Die Zeitspanne seit der Entstehung der Hominoidea. Basale Mitglieder eines Taxons können wenig Ähnlichkeit mit heutigen Formen haben, weil sie oft zahlreiche primitive Merkmale beibehalten. So könnte ein sehr früher Affe beispielsweise pronograd gewesen sein, auch wenn der letzte gemeinsame Vorfahre der lebenden Affen Anpassungen für eine aufrechte Haltung entwickelt hatte. Mögliche Synapomorphien zwischen Stamm- und Scheitelhominoiden dürften daher gering sein.
3) Die langen Evolutionsperioden in den existierenden Abstammungslinien könnten individuell einzigartig gewesen sein, so dass moderne Taxa nur wenige definierende Synapomorphien aufweisen, oder einige ihrer vermuteten Synapomorphien könnten Parallelismen sein.
4) Die Evolutionsgeschichte der morphologischen Veränderungen wird möglicherweise nicht angemessen geklärt, was zu widersprüchlichen phylogenetischen Interpretationen führt.
Vergleichende Studien, die sowohl Stamm- als auch Kronen-Katarrhinen einbeziehen, können diese Probleme entschärfen (Zalmout et al., 2010). So sind beispielsweise Merkmale, die bei einer spätoligozänen oder frühmiozänen Art vorhanden sind und die nicht primitiv mit Stammkatarrhinen und Cercopithecoiden geteilt werden, aber bei Kronenhominoiden beobachtet werden, plausible hominoide Synapomorphien. Zum Beispiel kommt der ethmo-frontale Sinus bei afrikanischen Menschenaffen vor und fehlt bei afro-arabischen Stammkatarrhinen, Cercopithecoiden, Pongo und Hylobates (Rossie et al., 2002; Rossie, 2008). Der Sinus kommt auch bei drei mutmaßlichen Hominoiden aus dem frühen Miozän vor: Proconsul, Morotopithecus und Afropithecus (ebd.). Die Verbreitung dieses Merkmals kann daher als ein abgeleitetes Merkmal interpretiert werden, das zuerst bei den Stammhominoiden auftrat und bei den asiatischen Affen verloren ging (Begun, 1992; Moyà-Solà und Köhler, 1995; siehe jedoch Rossie, 2008, für alternative Interpretationsmöglichkeiten dieses Merkmals). Ein weiteres Schädelmerkmal, die erhöhte Querschnittsstärke der Eckzähne und die damit einhergehende Vergrößerung des vorderen Gaumens, weist eine ähnliche Verteilung auf (d. h., es findet sich bei den heutigen Hominiden (beim Menschen sekundär verloren) sowie bei Proconsul, Morotopithecus und Afropithecus, fehlt aber bei den Stammkatarrhinen und Cercopithecoiden) und unterstützt ebenfalls die Interpretation, dass es sich bei den miozänen Taxa um Hominoide handelt (Begun et al., 1997; Zalmout et al., 2010). Der Kamoyapithecus aus dem späten Oligozän ist nur aus gnathodentalen Überresten bekannt (Leakey et al., 1995), aber das Vorhandensein von im Querschnitt erweiterten oberen Eckzähnen könnte darauf hindeuten, dass es sich ebenfalls um einen Hominoiden handelt (Zalmout et al., 2010). Darüber hinaus umfassen diese vier fossilen Gattungen Arten mit einer geschätzten Körpergröße von über 30 kg, die ersten Catarrhinen, die diese Größe erreichen.
Angesichts der Größe und der arborealen Nische der heute lebenden Menschenaffen (Gibbons sind wahrscheinlich sekundär zwergwüchsig: Tyler, 1993), könnte die große Körpergröße auch ein Merkmal der Stammhominoiden sein. Bei den Kronenhominoiden wird dieses Merkmal traditionell mit der Ausbeutung hochwertiger Baumnahrung durch die Hominoiden in Verbindung gebracht, die durch Innovationen im Bereich der Fortbewegung ermöglicht wurde, darunter die verbesserte Beweglichkeit der Gliedmaßen und die Fähigkeit zu greifen (wodurch das Gewicht auf mehrere Stützen verteilt werden kann) sowie die Orthogradie (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin und Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Belege für diese postkranialen Transformationen sind bei Proconsul, Afropithecus und Morotopithecus unterschiedlich ausgeprägt. Proconsul ist von mehreren Teilskeletten bekannt und wurde als Vierfüßer mit einem pronograden Rumpf rekonstruiert, dem jedoch ein Schwanz zu fehlen scheint (Napier und Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). Man rekonstruiert auch, dass Proconsul im Vergleich zu den primitiven Catarrhinen und Cercopithecoiden eine bessere Greiffähigkeit und eine größere Beweglichkeit in Gelenken wie dem Ellbogen besaß, was durch Merkmale wie eine ausgeprägte Zona conoidea im distalen Oberarmknochen belegt wird (Rose, 1988). Afropithecus ist postkranial weniger gut vertreten, ähnelt aber in den bekannten Elementen Proconsul (Ward, 1998). Obwohl der Zusammenhang zwischen den Schädel- und Postkranialproben aus den Moroto-Lokalitäten umstritten ist (Senut et al., 2000), unterscheiden sich die postkranialen Überreste von Morotopithecus durchweg von denen, die Proconsul zugeschrieben werden, und deuten stattdessen auf einen Hominoiden mit einem steifen unteren Rücken, einem beweglichen Schultergelenk und einem Oberschenkelknochen hin, der für absichtliches Klettern angepasst ist und eher an Kronenhominoide erinnert (MacLatchy et al, 2000).
Zusätzlich zur größeren Körpergröße weisen alle Taxa, die bewertet werden können, einen ausgeprägten Sexualdimorphismus sowohl bei der Körpergröße als auch bei der Morphologie der Eckzähne auf (Kelley, 1986; Ruff et al., 1989). Während der starke Sexualdimorphismus ein primitives Merkmal der Katarrhinen ist, ist er bei den scheinbaren Stammhominoiden und den modernen Menschenaffen weiter ausgeprägt (beim Menschen jedoch nur sekundär).
Es gibt nur wenige, wenn überhaupt, zwingende Synapomorphien im Gebiss der Hominoiden. Da die Hominoiden eine primitive katarrhine Zahnmorphologie beibehalten (Kay und Ungar, 1997), während die Zähne der Cercopithecoiden die eindeutige Synapomorphie der Bilophodontie der Backenzähne aufweisen, werden seit jeher schwach diagnostische Zahnmerkmale verwendet, um anzuerkennen, dass es sich bei einigen schlecht repräsentierten Katarrhinen des Oligozäns und Miozäns um Hominoiden handeln könnte, bis vollständigere Exemplare gefunden werden (z. B. Andrews, 1978 und 1992; Stevens et al., 2013). Solche oligozänen und miozänen Katarrhinen wurden informell als „Zahnaffen“ bezeichnet (z. B. Fleagle und Kay, 1987; Cartmill und Smith, 2009) oder unter einer Bezeichnung wie „Superfamily incertae sedis“ in die taxonomische Schwebe gestellt (z. B. Begun, 2007).
Es wurde die Idee geäußert, dass keines der oben diskutierten fossilen Taxa ein Hominoid ist und dass der erste erkennbare Hominoid der 14 Millionen Jahre alte Kenyapithecus ist, der einige abgeleitete kraniodentale Merkmale (z. B. ein reduziertes Molaren-Cingulum, eine höhere Position des Jochbogens) im Vergleich zu frühen miozänen Formen aufweist (Harrison, 2010). Sollte dies der Fall sein, würde zwischen dem geschätzten Ursprung der Hominoiden und ihrem ersten Auftreten eine unerklärliche Lücke von mindestens 10 Millionen Jahren klaffen – trotz eines robusten Fossilnachweises für einen Großteil dieses Zeitraums.
Obwohl Paläoanthropologen nicht in der Lage sind, die stammesgeschichtliche Stellung von „Zahnaffen“ oder Kamoyapithecus mit Sicherheit zu bestimmen, gibt es breite Unterstützung für die Hypothese, dass Proconsul, Afropithecus und Morotopithecus eher Stammhominoide als Stammkatarrhinen sind. Als Fenster in die frühe Hominoiden-Evolution legen diese Taxa Folgendes nahe:
1) Die große Körpergröße, die Vergrößerung des vorderen Gebisses und die subtilen Veränderungen im hinteren Gebiss (z. B. länglichere Backenzähne und Veränderungen in der Anordnung der Backenzähne und der Entwicklung des Zingulums) deuten darauf hin, dass sie grundlegend andere ökologische Nischen besetzen als die Stammkatarrhinen. Große Eckzähne dienen sowohl der Nahrungsbeschaffung als auch der sexuellen Selektion, wobei ein ausgeprägter Dimorphismus darauf hindeutet, dass Letzteres ein Faktor war. Eine große Größe ermöglicht den Zugang zu neuer Nahrung und verbessert die Flexibilität bei der Nahrungssuche (Wheatley, 1987), verringert das Raubtierrisiko, fördert das Überleben und kann in Verbindung mit Faktoren wie der Stabilität des Lebensraums mit einer verzögerten Reifung zusammenhängen. Die Analyse des Zahnentwicklungsmusters von Afropithecus (und möglicherweise Proconsul) deutet auf eine verzögerte Reifung im Vergleich zu anderen Anthropoiden hin (Kelley, 1997, 2002), was eine langsame Lebensgeschichte zu einer weiteren möglichen Stammhominoiden-Synapomorphie macht.
2) Bis zum frühen Miozän gibt es zumindest eine bescheidene Vielfalt an Fortbewegungs- und Nahrungsmitteln unter den großwüchsigen wahrscheinlichen Stammhominoiden, was auf die Anfänge einer echten adaptiven Radiation hindeutet, bei der sich gleichzeitig eine zunehmende Körpergröße, eine zunehmende Neuartigkeit der Fortbewegung und eine spezialisiertere Ausbeutung von Baumnahrungsquellen entwickeln.
3) Es ist derzeit nicht zu klären, welche anatomischen Merkmale, die mit Orthogradie und Gelenkbeweglichkeit bei modernen Menschenaffen verbunden sind, homolog oder homoplastisch sind (z.B. Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Diese Debatte hat Auswirkungen auf die Interpretation der evolutionären Position der angeblichen Stammhominoiden und auf die Rekonstruktion des Musters und des Zeitpunkts der Entstehung moderner Affenanpassungen.
So könnten beispielsweise postkraniale Ähnlichkeiten zwischen Morotopithecus und lebenden Affen auf verschiedene Weise interpretiert werden. Wenn die lebenden Menschenaffen ihre Orthogradie durch parallele Evolution erworben haben, dann stellt Morotopithecus das früheste bekannte Beispiel für einen Übergang dar, der in der Evolution der Hominoiden mehrmals stattfand. Dies ist von großem adaptivem Interesse, da es darauf hindeutet, dass eine zugrunde liegende entwicklungsbedingte oder genetische Neigung (siehe unten) die evolutionäre Reaktion der Hominoiden immer wieder in dieselbe Richtung lenkte. Dies würde auch bedeuten, dass bestimmte postkraniale Merkmale bei phylogenetischen Analysen mit Vorsicht zu genießen sind (Sanders und Bodenbender, 1994). Andererseits wäre Morotopithecus, wenn diese Ähnlichkeiten im Bewegungsapparat von einem gemeinsamen Vorfahren vererbt wurden, enger mit den Kronenhominoiden verwandt als Proconsul (und möglicherweise Afropithecus). Eine andere Möglichkeit ist, dass Morotopithecus ein unabhängiges „Experiment“ war und dass der gemeinsame Vorfahre der Kronenhominoiden dennoch orthograd war.
4) Wie oben erörtert, haben alle überlebenden Hominoiden-Taxa trotz der Unterschiede in der Bewegungsspezialisierung und der langen Perioden unabhängiger Evolution einen ähnlichen Körperbau, der sie von anderen Primaten unterscheidet (mit Ausnahme einiger Meerkatzen, deren Torso und Vordergliedmaßen sich konvergent zu denen der Affen entwickelt haben (Erikson, 1963; Larson, 1998). Die grundlegenden anatomischen Voraussetzungen des Körperbaus der Affen haben zu einer bemerkenswerten Vielfalt und Vielseitigkeit des Bewegungsapparats geführt, die es den großwüchsigen Primaten ermöglicht, sich effektiv in Bäumen (durch Klettern und Aufhängen) und auf der Erde (durch Knöchelgang und Zweibeinigkeit) fortzubewegen. Derartige Innovationen in der Fortbewegung, die mit Merkmalen wie der Orthogradie zusammenhängen, sind wahrscheinlich die Grundlage für den evolutionären „Erfolg“ aller lebenden Hominoiden und scheinen bis zu einem gewissen Grad auch bei den Stammhominoiden vorhanden gewesen zu sein. Entwicklungsarbeiten von Young und Kollegen (2010) haben gezeigt, dass die Integration zwischen Vorder- und Hintergliedmaßen bei Menschenaffen im Vergleich zu vierfüßigen Affen reduziert ist. Die Lockerung dieser genetischen Einschränkung wurde dahingehend interpretiert, dass sich die Gliedmaßen als Reaktion auf den Selektionsdruck freier auf unterschiedliche Weise entwickeln konnten (ebd.). Eine unterschiedliche Nutzung der Gliedmaßen bei Morotopithecus (MacLatchy et al., 2013) und jüngeren Taxa aus dem Miozän (z. B. Moya Sola und Kohler, 1996; Moya Sola et al., 2004; Nakatsukasa und Kunimatsu, 2009) stützen vorläufig die Vermutung von Young und Kollegen (2010), dass die „Evolvierbarkeit“ der Gliedmaßen ein ursprüngliches Merkmal der Hominoiden sein könnte, aber es wird mehr fossiles Material und ein besseres Verständnis der Genetik benötigt, die diesem Merkmal zugrunde liegt.
Künftige Erkenntnisse über die Ursprünge der Hominoiden und die frühe Evolution werden sich aus den fortgesetzten Versuchen ergeben, die Grundlage möglicher hominoider Synapomorphien zu identifizieren und zu verstehen. Dies wiederum erfordert einen verbesserten Fossilnachweis sowohl für die Stamm- als auch für die Scheitelkatarrhinen sowie eine detailliertere anatomische, entwicklungsgeschichtliche und genetische Untersuchung der zu untersuchenden gemeinsamen Merkmale. Die integrierte Untersuchung fossiler Hominoiden in ihrem paläoökologischen Kontext kann uns auch helfen, besser zu verstehen, warum hominoide Merkmale einmal oder mehrmals ausgewählt wurden.