HormonausschüttungBearbeiten
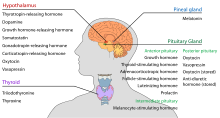
Der Hypothalamus hat eine zentrale neuroendokrine Funktion, vor allem durch seine Kontrolle der vorderen Hypophyse, die wiederum verschiedene endokrine Drüsen und Organe reguliert. Releasing-Hormone (auch Releasing-Faktoren genannt) werden in hypothalamischen Kernen produziert und dann entlang von Axonen entweder zur Eminenz oder zum Hypophysenvorderlappen transportiert, wo sie gespeichert und je nach Bedarf freigesetzt werden.
Hypophysenvorderlappen
In der Hypothalamus-Adenohypophysen-Achse werden die Releasing-Hormone, die auch als hypophysiotrope oder hypothalamische Hormone bezeichnet werden, von der medianen Eminenz, einer Verlängerung des Hypothalamus, in das hypophysäre Portalsystem freigesetzt, das sie zum Hypophysenvorderlappen transportiert, wo sie ihre regulierenden Funktionen auf die Sekretion der adenohypophysären Hormone ausüben. Diese hypophysiotropen Hormone werden durch parvozelluläre neurosekretorische Zellen stimuliert, die sich im periventrikulären Bereich des Hypothalamus befinden. Nach ihrer Freisetzung in die Kapillaren des dritten Ventrikels wandern die hypophysiotropen Hormone durch den so genannten hypothalamo-hypophysären Portalkreislauf. Sobald sie ihr Ziel im Hypophysenvorderlappen erreicht haben, binden sich diese Hormone an spezifische Rezeptoren, die sich auf der Oberfläche der Hypophysenzellen befinden. Je nachdem, welche Zellen durch diese Bindung aktiviert werden, beginnt die Hypophyse mit der Sekretion von Hormonen in den übrigen Blutkreislauf oder stellt diese ein.
| Ausgeschiedenes Hormon | Abkürzung | Produziert von | Wirkung |
|---|---|---|---|
| Thyreotropin-freisetzendes Hormon (Prolaktin-freisetzendes Hormon) |
TRH, TRF, oder PRH | Parvozelluläre neurosekretorische Zellen des Nucleus paraventricularis | Stimulieren die Freisetzung von Thyreoidea-stimulierendem Hormon (TSH) aus dem Hypophysenvorderlappen (hauptsächlich) Stimulieren die Freisetzung von Prolaktin aus dem Hypophysenvorderlappen |
| Corticotropin-Releasing Hormon | CRH oder CRF | Parvozelluläre neurosekretorische Zellen des Nucleus paraventricularis | Stimulieren die Freisetzung von adrenocorticotropem Hormon (ACTH) aus dem Hypophysenvorderlappen |
| Dopamin (Prolaktin-hemmendes Hormon) |
DA oder PIH | Dopamin-Neuronen des Nucleus arcuatus | Hindern die Prolaktinfreisetzung aus dem Hypophysenvorderlappen |
| Wachstums-hormonfreisetzendes Hormon | GHRH | Neuroendokrine Neuronen des Nucleus Arcuatus | Stimulieren die Freisetzung von Wachstums-Hormon (GH) aus dem Hypophysenvorderlappen |
| Gonadotropin-Releasing-Hormon | GnRH oder LHRH | Neuroendokrine Zellen des präoptischen Bereichs | Stimulieren Follikel-Stimulieren die Freisetzung des Follikel stimulierenden Hormons (FSH) aus dem Hypophysenvorderlappen Stimulieren die Freisetzung des luteinisierenden Hormons (LH) aus dem Hypophysenvorderlappen |
| Somatostatin (wachstumshormonhemmendes Hormon) |
SS, GHIH, oder SRIF | Neuroendokrine Zellen des Nucleus periventricularis | Hindern die Freisetzung von Wachstumshormon (GH) aus dem Hypophysenvorderlappen Hindern (mäßig) die Freisetzung von schilddrüsenstimulierendem Hormon (TSH) aus dem Hypophysenvorderlappen |
Zu den weiteren Hormonen, die von der Medianeminenz ausgeschüttet werden, gehören Vasopressin, Oxytocin und Neurotensin.
Hypophysenhinterlappen
In der Hypothalamus-Neurohypophysen-Achse werden neurohypophysäre Hormone aus dem Hypophysenhinterlappen, der eigentlich eine Verlängerung des Hypothalamus ist, in den Kreislauf abgegeben.
| Ausgeschiedenes Hormon | Abkürzung | Produziert von | Wirkung |
|---|---|---|---|
| Oxytocin | OXY oder OXT | Magnocelluläre neurosekretorische Zellen des Nucleus paraventricularis und des supraoptischen Nucleus | Uteruskontraktion Laktation (Absetzreflex) |
| Vasopressin (antidiuretisches Hormon) |
ADH oder AVP | Magnozelluläre und parvozelluläre neurosekretorische Zellen des Nucleus paraventricularis, magnozelluläre Zellen im Nucleus supraopticus | Erhöhung der Wasserdurchlässigkeit der Zellen des distalen Tubulus und des Sammelkanals in der Niere und damit Ermöglichung der Wasserrückresorption und der Ausscheidung von konzentriertem Urin |
Es ist auch bekannt, dass die Hormone der Hypothalamus-Hypophysen-Nebennieren-Achse (HPA) mit bestimmten Hautkrankheiten und der Hauthomöostase zusammenhängen. Es gibt Hinweise darauf, dass eine Hyperaktivität der HPA-Hormone mit stressbedingten Hauterkrankungen und Hauttumoren zusammenhängt.
StimulationBearbeiten
Der Hypothalamus koordiniert viele hormonelle und verhaltensbezogene zirkadiane Rhythmen, komplexe Muster neuroendokriner Leistungen, komplexe homöostatische Mechanismen und wichtige Verhaltensweisen. Der Hypothalamus muss daher auf viele verschiedene Signale reagieren, von denen einige extern und einige intern erzeugt werden. Delta-Wellen-Signale, die entweder im Thalamus oder in der Hirnrinde entstehen, beeinflussen die Sekretion von Releasing-Hormonen; GHRH und Prolaktin werden stimuliert, während TRH gehemmt wird.
Der Hypothalamus reagiert auf:
- Licht: Tageslänge und Photoperiode zur Regulierung zirkadianer und saisonaler Rhythmen
- Geruchsreize, einschließlich Pheromone
- Steroide, einschließlich gonadaler Steroide und Kortikosteroide
- Neural übertragene Informationen, die insbesondere vom Herzen, dem enterischen Nervensystem (des Magen-Darm-Trakts) und dem Fortpflanzungstrakt stammen.
- Autonome Eingänge
- Blutgebundene Reize, einschließlich Leptin, Ghrelin, Angiotensin, Insulin, Hypophysenhormone, Zytokine, Plasmakonzentrationen von Glukose und Osmolarität usw.
- Stress
- Eindringen von Mikroorganismen durch Erhöhung der Körpertemperatur, wodurch der Thermostat des Körpers nach oben versetzt wird.
Olfaktorische ReizeBearbeiten
Olfaktorische Reize sind für die sexuelle Fortpflanzung und die neuroendokrine Funktion bei vielen Arten wichtig. Wenn zum Beispiel eine trächtige Maus in einer kritischen Phase nach dem Koitus dem Urin eines „fremden“ Männchens ausgesetzt wird, scheitert die Schwangerschaft (Bruce-Effekt). So bildet ein Mäuseweibchen während des Koitus ein präzises „Geruchsgedächtnis“ für ihren Partner, das mehrere Tage lang bestehen bleibt. Pheromonale Hinweise helfen bei vielen Tierarten bei der Synchronisierung der Brunst; bei Frauen kann die synchronisierte Menstruation auch durch pheromonale Hinweise ausgelöst werden, obwohl die Rolle der Pheromone beim Menschen umstritten ist.
Stimuli aus dem BlutBearbeiten
Peptidhormone haben wichtige Einflüsse auf den Hypothalamus, und um dies zu tun, müssen sie die Blut-Hirn-Schranke passieren. Der Hypothalamus wird zum Teil von spezialisierten Hirnregionen begrenzt, die keine wirksame Blut-Hirn-Schranke besitzen; das Kapillarendothel an diesen Stellen ist gefenstert, so dass selbst große Proteine und andere Moleküle ungehindert passieren können. Einige dieser Stellen sind die Orte der Neurosekretion – die Neurohypophyse und die Eminenz mediana. Andere wiederum sind Stellen, an denen das Gehirn die Zusammensetzung des Blutes prüft. Zwei dieser Stellen, das SFO (subfornical organ) und das OVLT (organum vasculosum of the lamina terminalis), sind so genannte zirkumventrikuläre Organe, in denen die Neuronen in engem Kontakt mit Blut und Liquor stehen. Diese Strukturen sind dicht vaskularisiert und enthalten osmorezeptive und natriumrezeptive Neuronen, die das Trinken, die Vasopressinfreisetzung, die Natriumausscheidung und den Natriumappetit steuern. Sie enthalten auch Neuronen mit Rezeptoren für Angiotensin, atrialen natriuretischen Faktor, Endothelin und Relaxin, die alle für die Regulierung des Flüssigkeits- und Elektrolythaushalts wichtig sind. Die Neuronen des OVLT und des SFO projizieren in den Nucleus supraopticus und den Nucleus paraventricularis sowie in präoptische hypothalamische Bereiche. Die zirkumventrikulären Organe können auch der Wirkungsort von Interleukinen sein, die über Wirkungen auf paraventrikuläre Neuronen sowohl Fieber als auch ACTH-Sekretion auslösen.
Es ist nicht klar, wie alle Peptide, die die hypothalamische Aktivität beeinflussen, den notwendigen Zugang erhalten. Im Falle von Prolaktin und Leptin gibt es Hinweise auf eine aktive Aufnahme am Plexus choroideus aus dem Blut in die Zerebrospinalflüssigkeit (Liquor). Einige Hypophysenhormone haben einen negativen Rückkopplungseffekt auf die hypothalamische Sekretion; so wirkt beispielsweise das Wachstumshormon auf den Hypothalamus zurück, aber es ist nicht klar, wie es ins Gehirn gelangt. Es gibt auch Hinweise auf eine zentrale Wirkung von Prolaktin.
Es wurde festgestellt, dass das Schilddrüsenhormon (T4) von den hypothalamischen Gliazellen im Nucleus infundibularis/Medianus aufgenommen und hier durch die Typ-2-Deojodinase (D2) in T3 umgewandelt wird. Anschließend wird T3 in die Thyreotropin-Releasing-Hormon (TRH)-produzierenden Neuronen im Nucleus paraventricularis transportiert. In diesen Neuronen wurden Schilddrüsenhormonrezeptoren gefunden, was darauf hindeutet, dass sie tatsächlich empfindlich auf T3-Stimuli reagieren. Außerdem exprimierten diese Neuronen MCT8, einen Schilddrüsenhormon-Transporter, was die Theorie unterstützt, dass T3 in diese Neuronen transportiert wird. T3 könnte dann an den Schilddrüsenhormonrezeptor in diesen Neuronen binden und die Produktion des Thyreotropin-freisetzenden Hormons beeinflussen, wodurch die Schilddrüsenhormonproduktion reguliert wird.
Der Hypothalamus funktioniert als eine Art Thermostat für den Körper. Er legt eine gewünschte Körpertemperatur fest und stimuliert entweder die Wärmeproduktion und -speicherung, um die Bluttemperatur auf einen höheren Wert zu erhöhen, oder das Schwitzen und die Gefäßerweiterung, um das Blut auf eine niedrigere Temperatur abzukühlen. Alle Fieberzustände sind auf eine erhöhte Einstellung des Hypothalamus zurückzuführen; erhöhte Körpertemperaturen, die auf andere Ursachen zurückzuführen sind, werden als Hyperthermie eingestuft. In seltenen Fällen kann eine direkte Schädigung des Hypothalamus, z. B. durch einen Schlaganfall, Fieber verursachen; dies wird manchmal als hypothalamisches Fieber bezeichnet. Häufiger ist jedoch, dass eine solche Schädigung abnorm niedrige Körpertemperaturen verursacht.
SteroideEdit
Der Hypothalamus enthält Neuronen, die stark auf Steroide und Glukokortikoide reagieren – (die Steroidhormone der Nebenniere, die als Reaktion auf ACTH freigesetzt werden). Er enthält auch spezialisierte glukoseempfindliche Neuronen (im Nucleus arcuatus und im ventromedialen Hypothalamus), die für den Appetit wichtig sind. Der präoptische Bereich enthält wärmeempfindliche Neuronen, die für die TRH-Sekretion wichtig sind.
NeuralEdit
Die Oxytocin-Sekretion als Reaktion auf das Stillen oder die vagino-cervikale Stimulation wird durch einige dieser Bahnen vermittelt; die Vasopressin-Sekretion als Reaktion auf kardiovaskuläre Reize, die von Chemorezeptoren im Karotiskörper und im Aortenbogen sowie von Niederdruckrezeptoren des Vorhofvolumens ausgehen, wird durch andere vermittelt. Bei der Ratte bewirkt die Stimulation der Vagina auch eine Prolaktinsekretion, die nach einer unfruchtbaren Paarung zu einer Pseudoschwangerschaft führt. Beim Kaninchen löst der Koitus reflexartig einen Eisprung aus. Beim Schaf kann die Stimulation des Gebärmutterhalses in Anwesenheit hoher Östrogenspiegel bei einem jungfräulichen Mutterschaf mütterliches Verhalten hervorrufen. Alle diese Wirkungen werden vom Hypothalamus vermittelt, und die Informationen werden hauptsächlich über spinale Bahnen im Hirnstamm weitergeleitet. Die Stimulation der Brustwarzen regt die Freisetzung von Oxytocin und Prolaktin an und unterdrückt die Freisetzung von LH und FSH.
Kardiovaskuläre Reize werden durch den Vagusnerv weitergeleitet. Der Vagus überträgt auch eine Vielzahl viszeraler Informationen, darunter zum Beispiel Signale, die von der Magenerweiterung oder -entleerung ausgehen, um die Nahrungsaufnahme zu unterdrücken oder zu fördern, indem er die Freisetzung von Leptin bzw. Gastrin signalisiert. Auch diese Informationen erreichen den Hypothalamus über Relais im Hirnstamm.
Darüber hinaus reagiert die hypothalamische Funktion auf den Spiegel aller drei klassischen Monoamin-Neurotransmitter, Noradrenalin, Dopamin und Serotonin (5-Hydroxytryptamin), in den Bahnen, von denen sie innerviert wird, und wird von diesen reguliert. Zum Beispiel haben noradrenerge Eingänge, die vom Locus coeruleus ausgehen, wichtige regulatorische Auswirkungen auf den Spiegel des Corticotropin-Releasing-Hormons (CRH).
Kontrolle der NahrungsaufnahmeBearbeiten
| Peptide, die das Futter Verhalten |
Peptide, die das Fressverhalten verringern |
|---|---|
| Ghrelin | Leptin |
| Neuropeptid Y | (α,β,γ)-Melanozyten-stimulierende Hormone |
| Agouti-verwandtes Peptid | Kokain- und Amphetamin-regulierte Transkriptpeptide |
| Orexine (A,B) | Corticotropin-Releasing-Hormon |
| Melanin-konzentrierendes Hormon | Cholecystokinin |
| Galanin | Insulin |
| Glucagon-like peptide 1 |
Der äußerste seitliche Teil des ventromedialen Kerns des Hypothalamus ist für die Kontrolle der Nahrungsaufnahme verantwortlich. Die Stimulation dieses Bereichs bewirkt eine erhöhte Nahrungsaufnahme. Eine bilaterale Läsion dieses Bereichs führt zu einer vollständigen Einstellung der Nahrungsaufnahme. Mediale Teile des Nucleus haben eine kontrollierende Wirkung auf den lateralen Teil. Eine bilaterale Läsion des medialen Teils des Nucleus ventromedialis führt zu Hyperphagie und Fettleibigkeit bei den Tieren. Eine weitere Läsion des lateralen Teils des Nucleus ventromedialis bei demselben Tier führt zu einer vollständigen Einstellung der Nahrungsaufnahme.
Es gibt verschiedene Hypothesen im Zusammenhang mit dieser Regulation:
- Lipostatische Hypothese: Diese Hypothese besagt, dass das Fettgewebe ein humorales Signal produziert, das proportional zur Fettmenge ist und auf den Hypothalamus wirkt, um die Nahrungsaufnahme zu verringern und die Energieabgabe zu erhöhen. Es ist erwiesen, dass das Hormon Leptin auf den Hypothalamus einwirkt, um die Nahrungsaufnahme zu verringern und die Energieabgabe zu erhöhen.
- Gutpeptid-Hypothese: Gastrointestinale Hormone wie Grp, Glucagone, CCK und andere sollen die Nahrungsaufnahme hemmen. Die Nahrung, die in den Magen-Darm-Trakt gelangt, löst die Freisetzung dieser Hormone aus, die auf das Gehirn wirken, um Sättigung zu erzeugen. Das Gehirn enthält sowohl CCK-A- als auch CCK-B-Rezeptoren.
- Glucostatische Hypothese: Die Aktivität des Sättigungszentrums in den ventromedialen Kernen wird wahrscheinlich durch die Glukoseverwertung in den Neuronen gesteuert. Es wurde postuliert, dass die Aktivität in den Neuronen abnimmt, wenn ihre Glukoseverwertung niedrig ist und folglich die arteriovenöse Blutglukosedifferenz in ihnen gering ist. Unter diesen Bedingungen ist die Aktivität des Fütterungszentrums unkontrolliert und die Person fühlt sich hungrig. Die Nahrungsaufnahme wird durch die intraventrikuläre Verabreichung von 2-Desoxyglukose rasch gesteigert, wodurch die Glukoseverwertung in den Zellen sinkt.
- Thermostatische Hypothese: Nach dieser Hypothese regt ein Absinken der Körpertemperatur unter einen bestimmten Sollwert den Appetit an, während ein Anstieg über den Sollwert den Appetit hemmt.
AngstverarbeitungBearbeiten
Die mediale Zone des Hypothalamus ist Teil eines Schaltkreises, der motivierte Verhaltensweisen wie das Abwehrverhalten steuert. Analysen der Fos-Markierung haben gezeigt, dass eine Reihe von Kernen in der „Verhaltenskontrollsäule“ bei der Regulierung der Ausprägung von angeborenem und konditioniertem Abwehrverhalten eine wichtige Rolle spielt.
Abwehrverhalten bei Raubtieren
Die Begegnung mit einem Raubtier (z. B. einer Katze) löst bei Labornagern Abwehrverhalten aus, auch wenn das Tier noch nie einer Katze ausgesetzt war. Im Hypothalamus führt diese Exposition zu einem Anstieg der Fos-markierten Zellen im vorderen Hypothalamuskern, im dorsomedialen Teil des ventromedialen Kerns und im ventrolateralen Teil des prämammillären Kerns (PMDvl). Der prämammilläre Kern spielt eine wichtige Rolle bei der Ausprägung des Abwehrverhaltens gegenüber einem Raubtier, da Läsionen in diesem Kern Abwehrverhalten wie Frieren und Flucht aufheben. Der PMD moduliert das Verteidigungsverhalten in anderen Situationen nicht, da Läsionen dieses Kerns nur minimale Auswirkungen auf die Werte für das Einfrieren nach einem Schock haben. Der PMD hat wichtige Verbindungen zum dorsalen periaqueduktalen Grau, einer wichtigen Struktur für den Ausdruck von Angst. Darüber hinaus zeigen die Tiere ein Verhalten zur Risikobewertung der Umgebung, das zuvor mit der Katze in Verbindung gebracht wurde. Die Analyse von Fos-markierten Zellen zeigte, dass der PMDvl die am stärksten aktivierte Struktur im Hypothalamus ist, und die Inaktivierung mit Muscimol vor der Exposition gegenüber der Umgebung hebt das Abwehrverhalten auf. Daher spielt der Hypothalamus, vor allem der PMDvl, eine wichtige Rolle bei der Ausprägung des angeborenen und konditionierten Abwehrverhaltens gegenüber einem Raubtier.
Soziale Niederlage
Auch der Hypothalamus spielt eine Rolle bei der sozialen Niederlage: Kerne in der medialen Zone werden bei einer Begegnung mit einem aggressiven Artgenossen ebenfalls mobilisiert. Das besiegte Tier weist einen Anstieg der Fos-Werte in geschlechtsdimorphen Strukturen auf, wie dem medialen präoptischen Kern, dem ventrolateralen Teil des ventromedialen Kerns und dem ventralen prämammilären Kern. Diese Strukturen sind für andere soziale Verhaltensweisen, wie Sexual- und Aggressionsverhalten, von Bedeutung. Darüber hinaus ist auch der Nucleus premammillaris mobilisiert, und zwar der dorsomediale Teil, nicht aber der ventrolaterale Teil. Läsionen in diesem Kern heben passives Verteidigungsverhalten wie das Einfrieren und die „Rückenlage“ auf.