Kuvaus prokaryoottien transkriptio- ja translaatioprosessista
Prokaryootit, joihin kuuluvat bakteerit ja arkeologiset eliöt, ovat enimmäkseen yksisoluisia organismeja, joista puuttuu määritelmällisesti kalvoihin sidotut ytimet ja muut organellit. Bakteerikromosomi on kovalenttisesti suljettu ympyrä, joka, toisin kuin eukaryoottiset kromosomit, ei ole järjestäytynyt histoniproteiinien ympärille. Solun keskiosaa, jossa prokaryoottinen DNA sijaitsee, kutsutaan nukleoidiksi. Lisäksi prokaryooteilla on usein runsaasti plasmideja, jotka ovat lyhyempiä pyöreitä DNA-molekyylejä, jotka saattavat sisältää vain yhden tai muutaman geenin. Plasmidit voivat siirtyä bakteerikromosomista riippumatta solun jakautuessa, ja ne kantavat usein ominaisuuksia, kuten antibioottiresistenssiä. Näiden ainutlaatuisten ominaisuuksien vuoksi transkriptio ja geenien säätely eroavat jonkin verran prokaryoottisten solujen ja eukaryoottisten solujen välillä.
Oppimistavoitteet
- Ymmärtää DNA:n transkription perusvaiheet RNA:ksi prokaryoottisoluissa
- Ymmärtää prokaryoottisen translaation perusteet. ja miten se eroaa eukaryoottisesta translaatiosta
Prokaryoottinen transkriptio
Transkription käynnistyminen prokaryooteissa
Prokaryooteilla ei ole membraanisia-koteloituja tumia. Siksi transkription, translaation ja mRNA:n hajoamisen prosessit voivat tapahtua samanaikaisesti. Bakteeriproteiinin solunsisäinen taso voi nopeasti moninkertaistua useilla transkriptio- ja translaatiotapahtumilla, jotka tapahtuvat samanaikaisesti samalla DNA-mallilla. Prokaryoottinen transkriptio kattaa usein useamman kuin yhden geenin ja tuottaa polykistronisia mRNA:ita, jotka spesifioivat useamman kuin yhden proteiinin.
Keskustelemme tässä transkriptiota esimerkinomaisesti kuvaamalla tätä prosessia Escherichia coli -bakteerissa, joka on hyvin tutkittu bakteerilaji. Vaikka E. colin transkription ja arkeoiden transkription välillä on joitakin eroja, E. colin transkription ymmärtämistä voidaan soveltaa käytännöllisesti katsoen kaikkiin bakteerilajeihin.
Prokaryoottinen RNA-polymeraasi
Prokaryootit käyttävät samaa RNA-polymeraasia kaikkien geeniensä transkriptioon. E. colissa polymeraasi koostuu viidestä polypeptidi-alayksiköstä, joista kaksi on identtisiä. Neljä näistä alayksiköistä, jotka ovat α, α, β ja β′, muodostavat polymeraasin ydinentsyymin. Nämä alayksiköt kokoontuvat aina, kun geeni transkriboidaan, ja ne hajoavat, kun transkriptio on päättynyt. Kullakin alayksiköllä on oma tehtävänsä; kaksi α-alayksikköä ovat välttämättömiä polymeraasin kokoamiseksi DNA:lle; β-alayksikkö sitoutuu ribonukleosiditrifosfaattiin, josta tulee osa ”äskettäin syntynyttä” mRNA-molekyyliä; ja β′ sitoutuu DNA:n templaattijuosteeseen. Viides alayksikkö, σ, osallistuu vain transkription käynnistymiseen. Se antaa transkriptiospesifisyyden siten, että polymeraasi alkaa syntetisoida mRNA:ta sopivasta aloituskohdasta. Ilman σ:tä ydinentsyymi transkriboisi satunnaisista kohdista ja tuottaisi mRNA-molekyylejä, jotka spesifioivat proteiinien siansaksaa. Kaikista viidestä alayksiköstä koostuvaa polymeraasia kutsutaan holoentsyymiksi (holoentsyymi on biokemiallisesti aktiivinen yhdiste, joka koostuu entsyymistä ja sen koentsyymistä).
Prokaryoottiset promoottorit

Kuva 1. Prokaryoottisen RNA-polymeraasin σ-alayksikkö tunnistaa konsensussekvenssejä, jotka löytyvät promoottorin alueelta transkription aloituskohdasta ylävirtaan. σ-alayksikkö dissosioituu polymeraasista transkription käynnistyttyä.
Promoottori on DNA-sekvenssi, johon transkriptiokoneisto sitoutuu ja aloittaa transkription. Useimmissa tapauksissa promoottorit ovat niiden säätelemien geenien yläjuoksulla. Promoottorin erityinen sekvenssi on hyvin tärkeä, koska se määrää, transkriboidaanko vastaavaa geeniä koko ajan, jonkin verran vai harvoin. Vaikka promoottorit vaihtelevat eri prokaryoottigenomien välillä, muutamat elementit ovat konservoituneita. Aloituskohdasta ylävirtaan sijaitsevilla -10- ja -35-alueilla on kaksi promoottorin konsensussekvenssiä eli aluetta, jotka ovat samankaltaisia kaikissa promoottoreissa ja eri bakteerilajeissa (kuva 1).
-10-konsensussekvenssi, jota kutsutaan -10-alueeksi, on TATAAT. -35-sekvenssin, TTGACA, tunnistaa ja sitoo σ. Kun tämä vuorovaikutus on tapahtunut, ydinentsyymin alayksiköt sitoutuvat kohtaan. A-T-rikas -10-alue helpottaa DNA-mallin purkautumista, ja useita fosfodiesterisidoksia syntyy. Transkription initiaatiovaihe päättyy abortoivien transkriptien tuottamiseen, jotka ovat noin 10 nukleotidin pituisia polymeerejä, jotka valmistetaan ja vapautetaan.
Pidennys ja lopetus prokaryooteissa
Transkription pidennysvaihe alkaa σ- alayksikön irrottautumisella polymeraasista. σ:n dissosiaatio antaa ydinentsyymille mahdollisuuden edetä DNA-templaattia pitkin syntetisoiden mRNA:ta 5′-3′-suunnassa noin 40 nukleotidin sekuntinopeudella. Elongaation edetessä DNA purkautuu jatkuvasti ydinentsyymin edellä ja kelautuu uudelleen sen takana (kuva 2). DNA:n ja RNA:n välinen emäspariutuminen ei ole riittävän vakaata mRNA-synteesin komponenttien vakauden ylläpitämiseksi. Sen sijaan RNA-polymeraasi toimii vakaana linkkerinä DNA-mallin ja syntyvien RNA-juosteiden välillä varmistaakseen, ettei elongaatio keskeydy ennenaikaisesti.

Kuva 2. Klikkaamalla saat suuremman kuvan. Elongaation aikana prokaryoottinen RNA-polymeraasi seuraa DNA-mallia pitkin, syntetisoi mRNA:ta 5′:n ja 3′:n välisessä suunnassa ja purkaa ja kelaa DNA:ta takaisin sitä mukaa, kun sitä luetaan.
Prokaryoottiset lopetussignaalit
Kun geenin transkriptio on suoritettu, prokaryoottista polymeraasia on ohjeistettava irrottautumaan DNA-mallinaatista ja vapauttamaan äsken tuotettu mRNA. Transkriboitavasta geenistä riippuen on olemassa kahdenlaisia lopetussignaaleja. Toinen on proteiinipohjainen ja toinen RNA-pohjainen. Rho-riippuvaista terminaatiota ohjaa rho-proteiini, joka kulkee polymeraasin perässä kasvavassa mRNA-ketjussa. Lähellä geenin loppua polymeraasi törmää DNA-mallissa G-nukleotidien juoksuun, ja se pysähtyy. Tämän seurauksena rho-proteiini törmää polymeraasiin. Vuorovaikutus rhon kanssa vapauttaa mRNA:n transkriptiokuplasta.
Rho-riippumatonta terminaatiota kontrolloivat tietyt sekvenssit DNA-templaattijuosteessa. Kun polymeraasi lähestyy transkriboitavan geenin loppua, se kohtaa alueen, jossa on runsaasti C-G-nukleotideja. MRNA taittuu takaisin itseensä, ja komplementaariset C-G-nukleotidit sitoutuvat toisiinsa. Tuloksena on vakaa hiusnauha, joka saa polymeraasin pysähtymään heti, kun se alkaa transkriboida aluetta, jossa on runsaasti A-T-nukleotideja. mRNA-transkriptin komplementaarinen U-A-alue muodostaa vain heikon vuorovaikutuksen templaatti-DNA:n kanssa. Tämä yhdessä pysähtyneen polymeraasin kanssa saa aikaan riittävän epävakauden, jotta ydinentsyymi irtoaa ja vapauttaa uuden mRNA-transkriptin.
Transkription päättyessä transkriptioprosessi on valmis. Kun lopetus tapahtuu, prokaryoottinen transkripti olisi jo käytetty koodatun proteiinin lukuisten kopioiden synteesin aloittamiseen, koska nämä prosessit voivat tapahtua samanaikaisesti. Transkription, translaation ja jopa mRNA:n hajoamisen yhdistäminen on mahdollista, koska kaikki nämä prosessit tapahtuvat samassa 5′-3′-suunnassa ja koska prokaryoottisessa solussa ei ole kalvojen lokerointia (kuva 3). Sitä vastoin tuman läsnäolo eukaryoottisoluissa estää samanaikaisen transkription ja translaation.

Kuva 3. Useat polymeraasit voivat transkriboida yhtä bakteerigeeniä, kun taas lukuisat ribosomit kääntävät samanaikaisesti mRNA-transkriptioita polypeptideiksi. Näin tietty proteiini voi nopeasti saavuttaa suuren pitoisuuden bakteerisolussa.
Katso tämä BioStudio-animaatio nähdäksesi prokaryoottisen transkription prosessin.
Harjoituskysymykset
Minkä alayksikön E. coli-polymeraasi antaa transkription spesifisyyden?
- α
- β
- β′
- σ
Prokaryoottisten promoottorien -10- ja -35-alueita kutsutaan konsensusjaksoiksi, koska ________.
- ne ovat samanlaisia kaikissa bakteerilajeissa
- ne ovat samankaltaisia kaikissa bakteerilajeissa
- ne ovat olemassa kaikissa eliöissä
- neillä on sama tehtävä kaikissa eliöissä
Prokaryoottinen translaatio
Translaatio on samanlaista prokaryooteissa ja eukaryooteissa. Tässä tutkimme, miten translaatio tapahtuu E. coli -bakteerissa, joka on edustava prokaryootti, ja tarkennamme mahdolliset erot bakteeri- ja eukaryoottitranslaation välillä.
Initiaatio
Proteiinisynteesin käynnistyminen alkaa initiaatiokompleksin muodostumisella. E. colissa tähän kompleksiin kuuluu pieni 30S-ribosomi, mRNA-templaatti, kolme initiaatiotekijää, jotka auttavat ribosomia kasaantumaan oikein, guanosiinitrifosfaatti (GTP), joka toimii energianlähteenä, ja erityinen initiaattori-tRNA, joka kuljettaa N-formyylimetioniinia (fMet-tRNAfMet) (kuva 4). Initiaattori-tRNA on vuorovaikutuksessa mRNA:n aloituskodonin AUG kanssa ja kuljettaa formyloitua metioniinia (fMet). Koska fMet osallistuu initiaatioon, se lisätään jokaisen E. colin syntetisoiman polypeptidiketjun alkuun (N-terminaaliin). E. coli -mRNA:ssa ensimmäisen AUG-kodonin yläpuolella oleva johtava sekvenssi, jota kutsutaan Shine-Dalgarno-sekvenssiksi (tunnetaan myös nimellä ribosomaalinen sitoutumiskohta AGGAGAGG), on vuorovaikutuksessa komplementaarisen emäsparin kautta ribosomin muodostavien rRNA-molekyylien kanssa. Tämä vuorovaikutus ankkuroi 30S-ribosomaalisen alayksikön oikeaan paikkaan mRNA-mallissa. Tässä vaiheessa 50S-ribosomaalinen alayksikkö sitoutuu sitten aloituskompleksiin muodostaen ehjän ribosomin.
Eukaryooteissa initiaatiokompleksin muodostuminen on samanlaista, seuraavin eroin:
- Initiaattori-tRNA on erilainen erikoistunut metioniinia kantava tRNA, nimeltään Met-tRNAi
- Eukaryoottien initiaatiokompleksi ei sitoudu mRNA:han Shine-Dalgarno-sekvenssin kohdalla, vaan se tunnistaa eukaryoottien mRNA:n 5′:n korkin, minkä jälkeen se seuraa mRNA:ta pitkin pitkin 5′:n ja 3′:n välisessä suunnassa, kunnes AUG-käynnistyskodonki tunnistetaan. Tässä vaiheessa 60S-alayksikkö sitoutuu Met-tRNAi:n, mRNA:n ja 40S-alayksikön muodostamaan kompleksiin.

Kuva 4. Translaatio bakteereissa alkaa initiaatiokompleksin muodostumisella, johon kuuluu pieni ribosomaalinen alayksikkö, mRNA, N-formyylimetioniinia kantava initiaattori-tRNA ja initiaatiotekijöitä. Sitten 50S-alayksikkö sitoutuu, jolloin muodostuu ehjä ribosomi.
Pidennys
Prokaryooteissa ja eukaryooteissa translaation pidennyksen perusteet ovat samat. E. colissa 50S-ribosomaalisen alayksikön sitoutuminen tuottamaan ehjää ribosomia muodostaa kolme toiminnallisesti tärkeää ribosomaalista kohtaa: A-kohta (aminoasyyli) sitoo saapuvia varattuja aminoasyyli-tRNA:ita. P-paikka (peptidyyli) sitoo ladattuja tRNA:ita, jotka kantavat aminohappoja, jotka ovat muodostaneet peptidisidoksia kasvavan polypeptidiketjun kanssa, mutta eivät ole vielä irtautuneet vastaavasta tRNA:sta. E-kohta (exit) vapauttaa dissosioituneet tRNA:t, jotta ne voidaan ladata uudelleen vapailla aminohapoilla. Tässä tRNA:iden kokoonpanolinjassa on yksi huomattava poikkeus: Aloituskompleksin muodostumisen aikana bakteerien fMet-tRNAfMet tai eukaryoottinen Met-tRNAi menee suoraan P-kohtaan menemättä ensin A-kohtaan, jolloin vapautuu vapaa A-kohta, joka on valmiina ottamaan vastaan AUG:n jälkeistä ensimmäistä koodonia vastaavan tRNA:n.
Pidennys etenee ribosomin yksittäisten koodonien liikkeillä, joita kutakin kutsutaan translokaatiotapahtumaksi. Jokaisen translokaatiotapahtuman aikana varatut tRNA:t saapuvat A-kohtaan, siirtyvät sitten P-kohtaan ja lopulta E-kohtaan poistumista varten. Ribosomaaliset liikkeet eli askeleet saavat alkunsa konformaatiomuutoksista, jotka vievät ribosomia eteenpäin kolmella emäksellä 3′-suunnassa. A-kohdan tRNA:han kiinnittyneen aminohapon aminoryhmän ja P-kohdan tRNA:han kiinnittyneen aminohapon karboksyyliryhmän välille muodostuu peptidisidoksia. Kunkin peptidisidoksen muodostumista katalysoi peptidyylitransferaasi, RNA-pohjainen ribotsyymi, joka on integroitu 50S-ribosomaaliseen alayksikköön. P-kohdan tRNA:han sitoutunut aminohappo liittyy myös kasvavaan polypeptidiketjuun. Kun ribosomi astuu mRNA:n yli, entinen P-kohdan tRNA siirtyy E-kohtaan, irtoaa aminohaposta ja poistuu. Useat pidennyksen aikana tapahtuvat vaiheet, kuten ladatun aminoasyyli-tRNA:n sitoutuminen A-kohtaan ja translokaatio, vaativat energiaa, joka on peräisin GTP:n hydrolyysistä, jota katalysoivat erityiset pidennystekijät. Hämmästyttävää kyllä, E. colin translaatiolaitteisto tarvitsee vain 0,05 sekuntia kunkin aminohapon lisäämiseen, mikä tarkoittaa, että 200 aminohapon proteiini voidaan kääntää vain 10 sekunnissa.
Terminaatio
Translaation päättyminen tapahtuu, kun kohdataan nonsense-kodoni (UAA, UAG tai UGA), jolle ei ole komplementaarista tRNA:ta. Kohdistuessaan A-kohtaan nämä nonsense-kodonit tunnistetaan prokaryooteissa ja eukaryooteissa vapautumistekijöissä, jotka johtavat siihen, että P-kohdan aminohappo irtoaa tRNA:sta, jolloin vastavalmistunut polypeptidi vapautuu. Pienet ja suuret ribosomaaliset alayksiköt dissosioituvat mRNA:sta ja toisistaan; ne rekrytoituvat lähes välittömästi toiseen translaation init iointikompleksiin.
Yhteenvetona voidaan todeta, että on olemassa useita keskeisiä piirteitä, jotka erottavat prokaryoottisen geenin ilmentymisen eukaryooteissa esiintyvästä. Nämä on havainnollistettu kuvassa 5 ja lueteltu taulukossa 1.
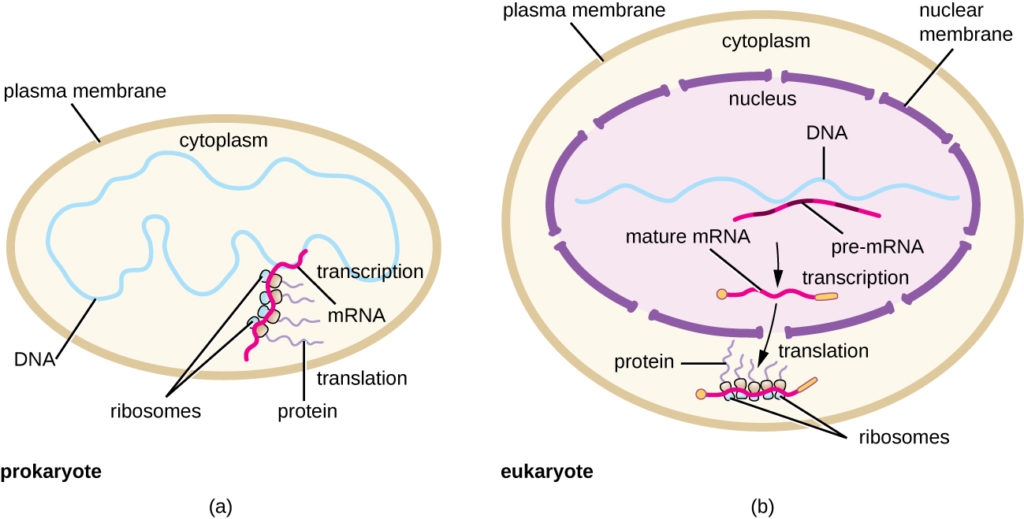
Kuva 5. (a) Prokaryooteissa transkriptio- ja translaatioprosessit tapahtuvat samanaikaisesti sytoplasmassa, mikä mahdollistaa solujen nopean reagoinnin ympäristön vihjeeseen. (b) Eukaryooteissa transkriptio on lokalisoitunut tumaan ja translaatio sytoplasmaan, mikä erottaa nämä prosessit toisistaan ja edellyttää RNA:n prosessointia stabiilisuuden vuoksi.
| Taulukko 1. Eukaryoottien transkriptio ja translaatio. Translation vertailu bakteereissa ja eukaryooteissa | ||
|---|---|---|
| Ominaisuus | Bakteerit | Eukaryootit |
| Ribosomit | 70S
|
80S
|
| Aminohappo, jota kuljettaa initiaattorin tRNA | fMet | Met |
| Shine-Dalgarno-sekvenssi mRNA:ssa | Esiintyy | Ei esiinny |
| Simultaaninen transkriptio ja translaatio | Kyllä | Ei |
Tarkista ymmärryksesi
Vastaamalla alla olevaan kysymykseen (kysymyksiin) näet, kuinka hyvin ymmärrät edellisessä osiossa käsitellyt aiheet. Tätä lyhyttä tietovisaa ei lasketa kurssin arvosanaan, ja voit suorittaa sen uudelleen rajoittamattoman määrän kertoja.
Käytä tätä tietovisaa tarkistaaksesi ymmärryksesi ja päättäessäsi, (1) opiskeletko edellistä osiota tarkemmin vai (2) siirrytkö seuraavaan osioon.