Yleisesittely
Molemmat meistä tietävät, että hiiva on erittäin hyödyllinen organismi erityisesti leivonnassa, viininvalmistuksessa ja panimossa. Mitä hiivat kuitenkin ovat ja miksi niitä tutkitaan niin paljon?

Hiivat ovat sieniä
Hiivat ovat yksisoluisia mikro-organismeja, jotka luokitellaan homeitiöiden ja sienien kanssa valtakunnan Sienet jäseniksi. Hiivat ovat evolutiivisesti monimuotoisia, ja siksi ne luokitellaan kahteen erilliseen heimoon, Ascomycota eli sakkasieniin ja Basidiomycota eli korkeampiin sieniin, jotka yhdessä muodostavat alaryhmän Dikarya. Nuppuhiivat, joita kutsutaan myös ”oikeiksi hiivoiksi”, kuuluvat heimoon Ascomycota ja järjestykseen Saccharomycetales. Tällaiset luokittelut perustuvat solun, ascospoorin ja pesäkkeen ominaisuuksiin sekä solufysiologiaan.
Hiivat ovat yksisoluisia, mutta niiden soluorganisaatio muistuttaa korkeampia organismeja
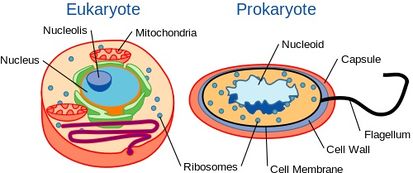
Vaikka hiivat ovat yksisoluisia organismeja, niillä on samanlainen soluorganisaatio kuin korkeammilla organismeilla, mukaan lukien ihminen. Erityisesti niiden geneettinen sisältö sijaitsee ytimessä. Tämä luokittelee ne eukaryoottisiksi organismeiksi, toisin kuin niiden yksisoluiset vastineet, bakteerit, joilla ei ole tumia ja joita pidetään prokaryootteina.
Luonnon elinympäristöt
Hiivat ovat luonnossa laajalti levinneet ja niillä on monenlaisia elinympäristöjä. Niitä tavataan yleisesti kasvien lehdissä, kukissa ja hedelmissä sekä maaperässä. Hiivoja esiintyy myös ihon pinnalla ja lämminveristen eläinten suolistossa, jossa ne voivat elää symbioottisesti tai loisina. Yleinen ”hiivatulehdus” on tyypillisesti Candida albicansin aiheuttama. Sen lisäksi, että Candida on emättimen hiivatulehdusten aiheuttaja, se aiheuttaa myös vaippaihottumaa sekä suun ja nielun sammasta.
Miksi tutkia hiivaa?
Kuvittele organismi, joka kasvaa nopeasti kolvissa ja jonka DNA:ta voidaan helposti manipuloida, mutta joka antaa myös tietoa ihmisen biologisista perusprosesseista, myös sairauksista. Hiiva sopii tähän kuvaukseen, ja se on tutkijoiden tutkimuskohteena kaikkialla maailmassa, ja tuloksena on yli 50 000 julkaistua tieteellistä artikkelia, joissa kuvataan hiivatutkimusta!

Mitkä hiivan erityispiirteet tekevät siitä ”malliorganismista”, jota tutkitaan ja johon kohdistuu niin paljon tutkimusta? Hiivat ovat yksisoluisia (yksisoluisia) organismeja, joten niitä on helppo tutkia, mutta niillä on samanlainen soluorganisaatio kuin korkeammilla, monisoluisilla organismeilla, kuten ihmisillä – eli niillä on tuma ja ne ovat siten eukaryootteja, kuten edellä kuvattiin. Tärkeintä on, että hiivan ja korkeampien eukaryoottien soluorganisaation samankaltaisuus johtaa samankaltaisuuteen niiden perustavanlaatuisissa soluprosesseissa, joten hiivassa tehdyt löydöt antavat usein suoria tai epäsuoria viitteitä siitä, miten biologiset prosessit toimivat ihmisessä.

Toinen tärkeä hiivojen ominaisuus, joka on olennainen niiden roolille ”malliorganismeina”, on se, että niitä on suhteellisen helppo käsitellä. Hiivat monistuvat nopeasti ja niitä on helppo manipuloida geneettisesti. Hiivan kaksinkertaistumisaika (aika, jonka solu tarvitsee monistumiseen ja jakautumiseen) on noin 90 minuuttia. Sitä vastoin viljelyssä kasvavat ihmissolut tarvitsevat noin 24 tuntia kaksinkertaistumiseen. Hiivalle on myös olemassa hyvin määriteltyjä geneettisiä menetelmiä, joiden avulla tutkijat voivat helposti eristää mutantteja, risteyttää niitä muiden mutanttien kanssa tai muihin geneettisiin taustoihin ja kartoittaa geenien sijainteja. Itse asiassa geenien välisen geneettisen etäisyyden perusteella rakennetut geneettiset kartat antoivat tutkijoille ensimmäisen näkemyksen genomista ja sen organisaatiosta, ja ne olivat 1900-luvun ensimmäiseltä puoliskolta peräisin olevien geneettisten tutkimusten huipentuma.
Keksintöjen kiihtynyt vauhti mahdollistui sen jälkeen, kun leivontahiivan (S. cerevisiae) genomista, joka edustaa sen täydellistä geneettistä materiaalia, tuli ensimmäinen eukaryoottien perimän sekvensointi jo vuonna 1996. Se on pienempi ja tiiviimpi kuin ihmisen genomi (12 miljoonaa emäsparia ja ~6 000 geeniä, kun taas ihmisen genomi on 3 miljardia emäsparia ja ~20-25 000 proteiinia koodaavaa geeniä). Silti genomien vertailut osoittavat, että ~31 prosenttia hiivan geeneistä on hyvin samankaltaisia kuin ihmisen geenit ja että 20 prosentilla ihmisen sairausgeeneistä on vastineensa hiivassa. Lisäksi hiivasolut voivat olla joko haploideja (yksi kromosomisarja) tai diploideja (kaksi kromosomisarjaa). Koska haploideissa on vain yksi kopio kustakin geenistä ja DNA-juosteiden katkeaminen ja yhdistyminen (rekombinaatio) on tehokasta, on hyvin helppoa poistaa tietty geeni haploidista ja tarkkailla vaikutuksia soluun eli deletoituneen mutantin ”fenotyyppiä”. Diploidisten solujen avulla puolestaan on mahdollista tutkia välttämättömiä geenejä (kasvun ja elinkelpoisuuden kannalta välttämättömiä geenejä) poistamalla yksi geenikopio ja tekemällä hienovaraisia muutoksia toiseen kopioon. Genomisekvenssistä saatujen tietojen avulla on luotu laaja molekyylireagenssien ja genomin laajuisten kokoelmien työkalupakki, joka tarjoaa tutkijoille tehokkaita välineitä biologisten ongelmien tutkimiseen. Jos hiivageenin tiedetään olevan DNA-sekvenssiltään samankaltainen kuin ihmisen geeni, hiivalla tehdyt tutkimukset voivat antaa tehokkaita vihjeitä sukulaisgeenin tai sukulaisgeenien roolista ihmisessä. Siten hiivan solutoimintojen tutkimisen suhteellinen yksinkertaisuus yhdistettynä sen merkitykseen korkeampiin eliöihin tekee siitä erittäin tehokkaan ”malliorganismin” tutkimukseen.
Hiivan elämä ja solusyklit
Hiivat kasvavat tyypillisesti suvuttomasti nuppuuntumalla. Emosoluun (emosoluun) muodostuu pieni silmu, josta tulee tytärsolu, ja se suurenee kasvun jatkuessa. Tyttösolun kasvaessa emosolu monistuu ja erottelee sitten DNA:nsa. Tydin jakautuu ja siirtyy tytärsoluun. Kun silmu sisältää tuman ja saavuttaa tietyn koon, se irtoaa emosolusta. Solussa tapahtuvia tapahtumasarjoja, jotka johtavat monistumiseen ja jakautumiseen, kutsutaan solusykliksi. Solusykli koostuu neljästä eri vaiheesta (G1, S, G2 ja M), ja sitä säädellään samalla tavalla kuin suurempien eukaryoottien solusykliä. Niin kauan kuin saatavilla on riittävästi ravinteita, kuten sokeria, typpeä ja fosfaattia, hiivasolut jatkavat jakautumistaan suvuttomasti.
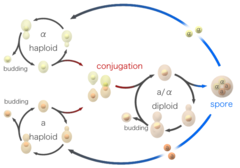
Hiivasolut voivat lisääntyä myös sukupuolisesti. Hiivasoluja esiintyy kahtena eri parittelutyyppinä, a-soluina ja alfasoluina. Kun vastakkaisia pariutumistyyppejä edustavia soluja sekoitetaan keskenään laboratoriossa tai ne joutuvat satunnaisesti kosketuksiin luonnossa, ne voivat pariutua (konjugoitua). Ennen liittymistä solut muuttavat muotoaan prosessissa, jota kutsutaan shmooingiksi. Termi ”shmoo” keksittiin, koska se muistuttaa muodoltaan Al Cappin 40-luvun lopulla luomaa samannimistä fiktiivistä sarjakuvahahmoa, joka esiintyi ensimmäisen kerran hänen sarjakuvassaan L’il Abner. Konjugaation aikana haploidiset solut sulautuvat ensin yhteen ja sitten niiden tumat, jolloin syntyy diploidinen solu, jossa on kaksi kappaletta kutakin kromosomia. Muodostuttuaan diploidiset solut voivat lisääntyä suvuttomasti nuppuuntumalla haploidien tapaan. Kun diploidiset solut kärsivät ravintoaineiden puutteesta, ne kuitenkin sporuloituvat. Sporulaation aikana diploidiset solut käyvät läpi meioosin, joka on erityinen solunjakautumisen muoto, jossa kromosomien määrä vähenee kahdesta kopiosta takaisin yhteen kopioon. Meioosin jälkeen meioosissa syntyneet haploidiset ytimet pakkautuvat neljäksi itiöksi, jotka sisältävät muokattuja soluseinämiä, jolloin syntyy rakenteita, jotka kestävät hyvin ympäristöstressiä. Nämä itiöt voivat säilyä hengissä pitkiä aikoja, kunnes olosuhteet muuttuvat suotuisammiksi, esimerkiksi paremmat ravinteet, jolloin ne pystyvät itämään ja lisääntymään suvuttomasti. Nämä eri tilat, orastuminen, konjugoituminen ja sporulaatio, muodostavat yhdessä hiivan elinkaaren.

Hiivan kasvu ja aineenvaihdunta
Kun hiivasoluja kasvatetaan runsaissa hiililähteissä, kuten glukoosissa, ne kasvavat mieluiten käymällä. Käymisen aikana glukoosi muuttuu hiilidioksidiksi ja etanoliksi. Yleensä käyminen tapahtuu ilman happea, joten se on luonteeltaan anaerobista. Jopa hapen läsnä ollessa hiivasolut kasvavat mieluummin fermentatiivisesti, ja tätä kutsutaan Crabtree-ilmiöksi biologin mukaan, joka löysi tämän preferenssin. Tätä kasvumuotoa hyödynnetään leivän, oluen, viinin ja muiden alkoholijuomien valmistuksessa. Vaikka orastavat hiivasolut kasvavat mieluiten käymisteitse, ravinteiden ollessa vähissä ne pystyvät kasvamaan myös soluhengityksen avulla. Hengityksen aikana solut muuttavat glukoosin hiilidioksidiksi ja vedeksi kuluttaen samalla happea, jolloin tuotetaan paljon suurempia määriä energiaa ATP:n muodossa.
Historialliset löydöt

Hiivaa on käytetty teollisena mikro-organismina jo tuhansia vuosia. Muinaiset egyptiläiset käyttivät hiivan käymistä leivän hapattamiseen. On todisteita jauhinkivistä, leivontakammioista ja piirroksia 4000 vuotta vanhoista leipomoista. Arkeologisissa kaivauksissa on löydetty todisteita 7000 vuotta vanhoja viinin jäänteitä sisältävien purkkien muodossa.
Mätiä visualisoi ensimmäisen kerran Antoni van Leeuwenhoek vuonna 1680 käyttämällä korkealaatuisia linssejä. Hän kuitenkin luuli, että nämä pallukat olivat tärkkelyspitoisia viljan hiukkasia, joita käytettiin vierteen, panimossa käytettävän nestemäisen uutteen, valmistukseen, eikä käymistilassa olevia hiivasoluja. Vuonna 1789 ranskalainen kemisti Antoine Lavoisier edisti ymmärrystämme niistä kemiallisista perusreaktioista, joita tarvitaan alkoholin tuottamiseen sokeriruo’osta. Arvioimalla lähtöaineiden ja tuotteiden (etanolin ja hiilidioksidin) osuudet hiivamassan lisäämisen jälkeen hän päätteli, että käytettiin kahta kemiallista reittiä, jossa kaksi kolmasosaa sokerista pelkistyi alkoholiksi ja yksi kolmasosa muodostui hiilidioksidiksi. Tuolloin kuitenkin ajateltiin, että hiiva oli vain mukana käynnistämässä reaktiota eikä sitä tarvittu koko prosessin ajan.
Vuonna 1815 ranskalainen kemisti Joseph-Louis Gay-Lussac kehitti menetelmiä viinirypälemehun pitämiseksi käymättömänä ja havaitsi, että käymättömän vierteen muuntamiseen tarvittiin ”fermentin” (joka sisälsi hiivaa) lisäämistä, mikä osoitti hiivan merkityksen alkoholikäymiselle. Vuonna 1835 Charles Cagniard de la Tour osoitti tehokkaamman mikroskoopin avulla, että hiivat olivat yksisoluisia ja lisääntyivät nuppujen avulla. Louis Pasteur havaitsi 1850-luvulla, että käymistilassa olevat juomat syntyivät, kun hiiva muutti glukoosin etanoliksi, ja määritteli käymisen ”hengitykseksi ilman ilmaa”. Lähes 1800-luvun lopulla Eduard Buchner käytti hiivasoluja jauhamalla saatuja soluttomia uutteita zymaasin, käymistä edistävien tai katalysoivien entsyymien kokoelman, havaitsemiseen, ja tästä hänelle myönnettiin Nobel-palkinto vuonna 1907.
Paljon hiivan genetiikan uraauurtavaa työtä teki Øjvind Winge. Hän havaitsi, että hiivat vuorottelevat haploidisten ja diploidisten tilojen välillä ja että hiivat ovat heterotallikoita, sillä haploidien muuttamiseen diploideiksi (konjugaatio) tarvitaan kaksi kantaa. Hän ja hänen kollegansa Otto Laustsen kehittivät tekniikoita hiivojen mikromanipuloimiseksi, jotta niitä voitaisiin tutkia geneettisesti. Tässä tekniikassa, joka tunnetaan nimellä ”tetrad-analyysi”, käytetään hienoa neulaa ja mikroskooppia eristämään rakenne, joka tunnetaan nimellä ascus ja joka sisältää neljä itiötuotetta eli tetradia, jotka ovat tulosta diploidin sporulaatiosta. Kun ascus on eristetty, tetradin itiöt erotetaan toisistaan ja niiden annetaan kasvaa pesäkkeiksi geneettistä analyysia varten. Tämä uraauurtava työ toi hänelle arvonimen ”hiivagenetiikan isä”. Osaa tästä työstä selvitti edelleen Carl Lindegren, joka selvitti parittelutyyppijärjestelmän orastuvassa hiivassa osoittamalla Mat a- ja Mat alfa-solujen olemassaolon, kehitti menetelmiä näiden parittelutyyppien solujen välisten massaparittelujen toteuttamiseksi ja käytti tätä tietoa sokerin hyväksikäytön genetiikan tutkimiseen.
Sen jälkeen moni muukin tutkija on tehnyt uraauurtavia tutkimuksia orastuvaa hiivaa käyttäen. Joillekin näistä tutkijoista on myönnetty Nobel-palkinto näiden tutkimusten aikana tehdyistä merkittävistä löydöistä, mm: Tohtori Leland Hartwell (2001) solusykliä säätelevien geenien löytämisestä (yhdessä Paul Nursen ja Tim Huntin kanssa); Roger Kornberg (2006) tutkimuksistaan, jotka koskivat geeniekspression ensimmäistä vaihetta, eli tapaa, jolla geenin DNA-sekvenssi kopioidaan sanansaattaja-RNA:ksi (mRNA:ksi); tohtorit Leland Hartwell (2001) ja Tim Hunt (2006). Elizabeth Blackburnille, Carol Greiderille ja Jack Szostakille (2009) geeneistä ja keinoista, joilla solut suojaavat kromosomien päitä eli telomeerejä hajoamiselta, ja tohtoreille Randy Schekmanille, James Rothmanille ja Thomas Südhofille (2013) vesikulaariliikennettä säätelevää koneistoa koskevista tutkimuksista. Viimeksi tohtori Yoshinori Ohsumi sai palkinnon autofagiaa koskevasta työstään, joka alkoi hiivalla tehdyistä tutkimuksista.
Kaupalliset sovellukset

Hiivaa on pitkään pidetty parhaana organismina alkoholijuomien, leivän ja monien erilaisten teollisuustuotteiden valmistuksessa. Tämä perustuu siihen, että hiivan aineenvaihduntaa voidaan helposti manipuloida geenitekniikoiden avulla, että sitä voidaan kasvattaa nopeasti suuriin solusatoihin (biomassaan), että tämä biomassa voidaan helposti erottaa tuotteista ja että se on yleisesti tunnustettu turvalliseksi (GRAS).
S. cerevisiae -hiivaa ja muita hiivalajeja on pitkään käytetty riisin, vehnän, ohran ja maissin sokerien käymiseen alkoholijuomien, kuten oluen ja viinin, valmistamiseksi. Panimohiivoja on kahta päätyyppiä, ylhäältä käyvää ale-hiivaa ja alhaalta käyvää lagerhiivaa. Yläkäyvä hiiva, kuten S. cerevisiae nousee pinnalle käymisen aikana, ja niitä käytetään alen, portterin, stoutin ja vehnäoluen valmistukseen. Sitä vastoin S. pastorianus (entinen S. carlsbergensis) on alhaalla käyvä hiiva, jota käytetään lager-oluen valmistukseen. Lagerhiivat kasvavat parhaiten matalammissa lämpötiloissa. Tämän seurauksena ne kasvavat hitaammin, tuottavat vähemmän pintavaahtoa ja laskeutuvat siksi tyypillisesti käymisastian pohjalle. Pilsnerit, Märzenit, Bockit ja amerikkalaiset mallasoluet ovat kaikki lageroluen tyylejä. Nykyaikaisessa panimotoiminnassa monet alkuperäisistä ylhäältä käyvistä kannoista on muunnettu alhaalta käyviksi.
Mehiläiset tuottavat viiniä käymällä rypäletäysmehun (rypäleen puristemehun) sokerit etanoliksi. Vaikka viinin käyminen voi käynnistyä viinitarhoissa luonnostaan esiintyvällä hiivalla, monet viinitilat päättävät lisätä puhdasta hiivaviljelmää käymisen hallitsemiseksi ja valvomiseksi. Samppanjan ja kuohuviinien kuplat syntyvät tyypillisesti pullossa tapahtuvassa jälkikäymisessä, joka sitoo hiilidioksidia. Viinin valmistuksessa syntyvä hiilidioksidi vapautuu sivutuotteena. Yksi hiivasolu voi käydä noin oman painonsa verran glukoosia tunnissa. Optimaalisissa olosuhteissa S. cerevisiae voi tuottaa jopa 18 tilavuusprosenttia etanolia, mutta tavallisesti etanolipitoisuus on 15-16 prosenttia. Kaupallisesti tuotetussa viinissä esiintyvä rikkidioksidi lisätään heti rypäleiden murskauksen jälkeen, jotta luonnostaan esiintyvät bakteerit, homeet ja hiivat saadaan tapettua.

Saccharomyces cerevisiae eli leivontahiiva on jo pitkään ollut leivonnassa käytetty hapatusaine. Leipurihiiva fermentoi taikinassa olevia sokereita tuottaen hiilidioksidia ja etanolia. Hiilidioksidi jää kiinni taikinan pieniin kupliin, mikä saa taikinan kohoamaan. Hapanjuurileipä on poikkeus, sillä sitä ei valmisteta leivontahiivalla vaan villihiivan ja bakteerien yhdistelmällä. Candida milleri -hiivaa käytetään gluteenin vahvistamiseen, ja happoa tuottavaa Lactobacillus sanfranciscensis -bakteeria käytetään maltoosin käymiseen.
Näiden perinteisten käyttötarkoitusten lisäksi hiivaa on käytetty myös moniin muihin kaupallisiin sovelluksiin. Vegaanit käyttävät hiivaa usein juuston korvikkeena ja sitä käytetään usein esimerkiksi popcornin kaltaisten tuotteiden täytteenä. Sitä käytetään petrokemianteollisuudessa, jossa sitä on kehitetty tuottamaan biopolttoaineita, kuten etanolia ja farneseenia, joka on dieselin ja lentopetrolin esiaste. Sitä käytetään myös voiteluaineiden ja pesuaineiden tuotannossa. Elintarviketeollisuudessa hiivaa käytetään elintarvikelisäaineiden, kuten väriaineiden, antioksidanttien ja arominvahventeiden valmistukseen. Sitä käytetään usein lääkkeiden, kuten parasiittilääkkeiden, syöpää ehkäisevien yhdisteiden, biofarmaseuttisten tuotteiden, kuten insuliinin, rokotteiden ja ravitsemuslääkkeiden valmistuksessa. Hiivaa käytetään yleisesti teollisten entsyymien ja kemikaalien tuotannossa. Ympäristön bioremediaation alalla kantoja on hyödynnetty jopa metallien poistamiseen kaivosjätteestä.
Sovellus ihmisen sairauksiin ja tutkimukseen

Hiivan geenien ja ihmisen vastaavien geenien suuren samankaltaisuuden ja konservoidun perustavanlaatuisen solubiologian ansiosta hiivasta on tullut suosittu mallisysteemi ihmisen sairausgeenien tutkimiseen. Kun ihmisen ja hiivan geenin välille on löydetty yhteys, on käytetty useita lähestymistapoja ihmisen geenien tuntemuksen lisäämiseksi. Eräässä lähestymistavassa ihmisen sairauteen liittyvän geenin löydyttyä sen sekvenssiä verrataan kaikkien hiivan genomissa olevien geenien sekvensseihin, jotta löydetään samankaltaisin hiivageeni tai samankaltaisimmat hiivageenit. Sen tutkimiseksi, ovatko geenit toiminnallisesti sukua toisilleen, ihmisen geeni ekspressoidaan sitten hiivatahrassa, jossa hiivageeni on ensin inaktivoitu mutaation avulla. Näin tutkijat voivat määrittää, pystyykö ihmisgeeni pelastamaan hiivageenin elinkelpoisuuden, kasvun tai muita hiivageenin menettämiseen liittyviä erityisiä vikoja, menetelmää kutsutaan toiminnalliseksi komplementoinniksi. Jos reitit ja/tai prosessit, joihin hiivageeni osallistuu, ovat konservoituneita, ihmisgeenin toiminnasta voidaan oppia paljon sen perusteella, mitä vastaavasta hiivageenistä jo tiedetään. Kun funktionaalinen komplementaatio on todettu, tutkijat voivat käyttää tätä järjestelmää kuvaamaan tarkemmin ihmisen geenituotteen toimintaa. Vähemmän kohdennetut lähestymistavat, joissa käytetään usein korkean läpimenon tekniikoita (HTP), joilla seulotaan satunnaisesti tuhansia ihmisen geenejä kerralla, jotta voidaan löytää geeni tai geenejä, joilla on komplementtiaktiivisuutta. Tällaisia lähestymistapoja on käytetty menestyksekkäästi konservoituneiden solusyklin säätelijöiden (CDC2), syöpään osallistuvien geenien ja hermoston rappeutumissairauksiin osallistuvien geenien tunnistamiseen.
On olemassa monia skenaarioita, joissa tehdyt tutkimukset voivat antaa arvokasta informaatiota tutkijoille siitä, millaisiin soluvirtausketjuihin ja/tai -prosesseihin ihmisgeenin geenin osallistuminen vaikuttaa, kun sukulaisgeeniä sisältävää hiivageeniä ei ole. Esimerkiksi jotkin hermoston rappeutumissairaudet, kuten Alzheimerin ja Parkinsonin tauti, syntyvät, kun amyloidiksi kutsutut proteiiniaggregaatit kerääntyvät proteiinien vääränlaisen taittumisen vuoksi, ja tämä on myrkyllistä hermosoluille. Tutkijat ovat saaneet tietoa näistä hermoston rappeutumissairauksista tutkimalla hiivaproteiineja, joilla on samanlainen amyloidinmuodostuspotentiaali ja joita kutsutaan prioneiksi. Vaihtoehtoisesti sairauteen liittyvän geenin lisääntynyt ilmentyminen hiivassa voi johtaa fenotyyppiin. Esimerkiksi Parkinsonin tautiin liittyvä alfa-synukleiini on myrkyllinen, kun sitä ilmentyy riittävän paljon. Tällaista kantaa voidaan sitten käyttää sellaisten hiivageenien tai pienten molekyylien seulontaan, jotka tukahduttavat tai tehostavat synukleiinin aiheuttamaa myrkyllisyyttä, jolloin saadaan usein vihjeitä asiaan liittyvistä solupoluista. Amyotrofista lateraaliskleroosia (ALS) tai Lou Gehrigin tautia sairastavilla potilailla on usein mutaatioita parissa RNA:ta sitovassa proteiinissa, mikä altistaa ne muodostamaan aggregaatteja, jotka häiritsevät RNA:n aineenvaihduntaa. Hiivaseulonnalla on onnistuttu tunnistamaan useita hiivageenejä, joilla on samankaltaisia ominaisuuksia (muodostavat myrkyllisiä aggregaatteja), ja näin tutkijat ovat saaneet uusia geeniehdokkaita tutkittavaksi. Vastaavasti ihmisen RNA:ta sitovat proteiinit muodostavat hiivassa ilmentyessään myrkyllisiä aggregaatteja, ja tätä kantaa käytettiin sellaisen hiivageenin tunnistamiseen, joka mutaationa estää näiden aggregaattien muodostumisen.
Hiivasta on tulossa ensisijainen organismi tutkimuksissa, joiden tarkoituksena on tunnistaa lääkkeiden kohteita ja erilaisten lääkkeiden vaikutustapaa. Kemogenomiikka tai kemiallis-genomiikka tarkoittaa seuloja, joissa käytetään kemikaalien ja genomiikan yhdistelmää lääkekohteiden tutkimiseen ja mahdollisesti uusien lääkkeiden tunnistamiseen. Näissä kemiallis-genomiikan tutkimuksissa on käytetty kahta päälähestymistapaa. Ensimmäisessä menetelmässä rakennetaan koko genomin kattava kokoelma diploidisia kantoja, joissa toinen geenin kahdesta identtisestä kopiosta on poistettu, mikä alentaa tietyn geenituotteen määrää. Kohdegeenit ja kohdegeenireittiin osallistuvat geenit tulevat herkemmiksi yhdisteelle, ja ne tunnistetaan ensisijaisesti tällaisessa seulonnassa. Toisessa lähestymistavassa epäolennaisia geenejä poistetaan systemaattisesti ja kokoelmaa seulotaan lääkkeellä, jotta voidaan etsiä geenejä, jotka puskuroivat lääkkeen kohdepolkua. Tämän lähestymistavan odotetaan tunnistavan geenejä, joita tarvitaan kasvuun yhdisteen läsnä ollessa. Muita lähestymistapoja, joissa käytetään yliekspressioseulontoja, on käytetty tunnistamaan lääkeresistenssiin osallistuvia geenejä, mukaan lukien potentiaalinen lääkekohde. Jonkin geenin osalta deletoitujen hiivasolujen ilmentymisprofiilin vertaaminen tietyllä lääkkeellä käsiteltyjen villityyppisten hiivasolujen ilmentymisprofiiliin voi myös olla tehokas tapa tunnistaa geenejä, jotka saattavat kertoa tutkijoille jotakin siitä, miten lääke toimii soluissa.
Nämä ovat vain muutamia esimerkkejä siitä, miten hiivaa voidaan käyttää sekä ihmisen sairauksien tutkimiseen. Hiivalla tehdyt tutkimukset voivat auttaa tutkijoita oppimaan enemmän taustalla olevasta biologiasta tämän mallijärjestelmän avulla tai auttaa heitä tunnistamaan lääkekohteita tai lääkkeiden vaikutustapaa.
Lähteet
- Saccharomyces-genomitietokanta (Saccharomyces Genome Database, SGD) tarjoaa kattavaa integroitua biologista tietoa orastuvasta hiivasta Saccharomyces cerevisiae sekä haku- ja analyysityökaluja, joilla voidaan tutkia näitä tietoja.
- MIPS Comprehensive Yeast Genome Database (CYPD) esittelee tietoa kokonaan sekvensoidun, hyvin tutkitun mallieukaryootin, orastavan hiivan Saccharomyces cerevisiae, molekyylirakenteesta ja toiminnallisesta verkostosta.
- Candida Genome Database (CGD), Candida albicansin genomin sekvenssidatan sekä geenejä ja proteiineja koskevan tiedon lähde.
- PomBase, kattava tietokanta fissiohiivalle Schizosaccharomyces pombe, joka tarjoaa rakenteellista ja funktionaalista annotaatiota, kirjallisuuden kuratointia ja pääsyn laajoihin tietokokonaisuuksiin.
- Fungal genome resources at NCBI, opas sienienien genomiresursseihin kansallisessa biotekniikkatietokeskuksessa (NCBI).
- Laaja luettelo hiivaan liittyvistä resursseista aiheista, jotka vaihtelevat yleisestä hiivatiedosta nukleiinihappoihin, genomeihin ja proteiineihin, ekspressiotietoihin, lokalisaatioon, fenotyyppeihin ja muuhun.
Suositeltava lukeminen
Kirjat
- YeastBook. (2011) Kattava katsausten kokoomateos, joka esittelee hiivan Saccharomyces cerevisiae molekyylibiologian, solubiologian ja genetiikan tämänhetkisen tietämyksen, Genetiikka
- From a to alpha: Hiiva solujen erilaistumisen mallina. (2007) Hiten D. Madhani, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Landmark Papers in Yeast Biology. (2006), toimittaneet Patrick Linder, David Shore ja Michael N. Hall, Cold Spring Harbor Press, Cold Spring Harbor, New York.
- Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual. (2005) David C. Amberg, Daniel J. Burke ja Jeffrey N. Strathern. Cold Spring Laboratory Press, Cold Spring Harbor, New York.
- The Early Days of Yeast Genetics. (1993) Toimittaneet Michael N. Hall ja Patrick Linder. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Volume I: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Genome Dynamics, Protein Synthesis, and Energetics. (1991) toimittaneet James R. Broach, John R. Pringle ja Elizabeth W. Jones. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Volume II: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Gene Expression. (1992) toimittaneet Elizabeth W. Jones, John R. Pringle ja James R. Broach. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Volume III: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Cell Cycle and Cell Biology. (1997) toimittaneet John R. Pringle, James R. Broach ja Elizabeth W. Jones. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Hiiva: A Practical Approach. (1988) Toimittaneet I. Campbell ja , ja John H. Duffus, IRL Press, Ithaca, New York.
Lehtiartikkelit
- Duina A.A., Miller M.E., ja J.B. Keeney (2014) Budding Yeast for Budding Geneticists: A Primer on the Saccharomyces cerevisiae Model System. Genetics. 197:33-48.
- Botstein D. ja G.R. Fink (2011) Yeast: An Experimental Organism for 21st Century Biology. Genetics. 189:695-704.
- Cherry J.M., Hong E.L., Amundsen C., Balakrishnan R., Binkley G., Chan E.T., Christie K.R., Costanzo M.C., Dwight S.S., Engel S.R., Fisk D.G., Hirschman J.E., Hitz B.C., Karra K., Krieger C.J., Miyasato S.R., Miyasato S.R., Nash R.S., Park J., Skrzypek M.S., Simison M., Weng S. ja E.D. Wong (2011) Saccharomyces Genome Database: the genomics resource of budding yeast. Nucleic Acids Res. 40:D700-D705.
- Altman T.J., Boone C., Churchill G.A., Hengartner M.O., Mackay T.F. ja D.L. Stemple (2011) Malliorganismien tulevaisuus ihmisen sairauksien tutkimuksessa. Nat. Rev. Genet. 18:575-582.
- Ho C.H., Piotrowski J., Dixon S.J., Baryshnikova A., Costanzo M. ja C. Boone (2011) Combining functional genomics and chemical biology to identify targets of bioactive compounds. Curr Opin Chem Biol. 15:66-78.
- Smith A.M., Ammar R., Nislow C. ja G. Giaever (2010) A survey of yeast genomic assays for drug and target discovery. Pharmacol Ther. 127:156-164.
- Dolinski K. ja D. Botstein (2007) Orthology and functional conservation in eukaryotes. Annu. Rev. Genet. 41:465-507.
- Barnett J.A. (2007) A history of research on yeasts 10: foundations of yeast genetics. Yeast 24:799-845.
- Spradling A., Ganetsky B., Hieter P., Johnston M., Olson M., Orr-Weaver T., Rossant J., Sanchez A. ja R. Waterston (2006) New roles for model genetic organisms in understanding and treating human disease: report from the 2006 Genetics Society of America meeting. Genetics 172:2025-2032.
- Sherman F. (2002) Getting started with yeast Methods Enzymol. 350:3-41.