T.H. Huxley azt írta, hogy más majmokra nézve “önmagunk elmosódott másolatait” látjuk (1863 pg. 73). A majmok, vagyis a hominoidák közötti szoros testi rokonságot már egy évszázaddal korábban felismerte Carolus Linnaeus (1759). Az élő hominoidákat a habituális ortogradia és az ágak alatti viselkedéssel kapcsolatos tulajdonságok kötik össze: széles törzs széles vállízületekkel, merev hát, hosszú mellső végtagok, mozgékony végtagízületek, erős kapaszkodóképesség és a farok hiánya (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Az emberből számos ilyen tulajdonság, például a viszonylag hosszú karok, a kétlábúsághoz való másodlagos alkalmazkodás eredményeként hiányzik. A koponyatulajdonságok is egyesítik a hominoidákat, köztük a széles elülső szájpadlás és a viszonylag nagy agy (Jerison, 1973; Rae, 1997). A hominoidák élettartama is meghosszabbodott, késleltetett éréssel (Kelley, 1997), és hasonló ökológiai fülkéket foglalnak el – az ember kivételével valamennyi faj trópusi erdőkben él, és táplálkozásuk jelentős mértékben az érett gyümölcsökre és levelekre támaszkodik (Fleagle, 2013).
A hylobatidák (gibbonok és sziámángok) kisebbek, mint a többi élő majom, és más szempontból is különböznek, ami a hominidáktól (emberszabású majmok és emberek), amelyeknek egy közelebbi közös ősük van, hosszú evolúciós elszigetelődésüket tükrözi (1. ábra). A hominidák (az ember kivételével) elülső szájpadlása a hylobatidákénál szélesebb (Rae, 1997), még rövidebb, merevebb alsó háttal rendelkeznek, és hiányzik a ricochetális brachiáció anatómiai specializációja (Young, 2003).
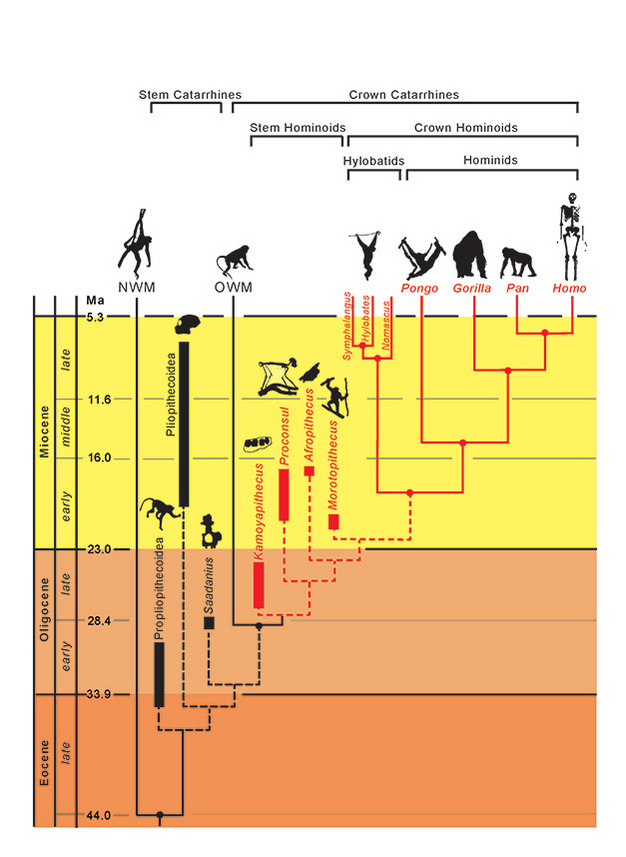
A taxonok filogenetikai kapcsolatait folytonos (kihalt) és szaggatott (kihalt) vonalak jelzik. A földtörténeti időt a függőleges tengelyen ábrázoljuk. A taxonok elágazási eseményeinek időzítése csak hozzávetőleges, és molekuláris és fosszilis adatok kombinációjából következtethető (Steiper et al., 2004; MacLatchy et al., 2006; Steiper és Young, 2008 és 2009; Chan et al., 2010; Stevens et al., 2013). A fosszilis taxonok filogenetikai elhelyezései hipotetikusak, amint azt a szövegben kifejtjük. Az Afropithecus és a Morotopithecus helyzete megfordulhat, vagy a kettő lehet testvértaxon is. Az NMW az újvilági majmokra, az OWM pedig az óvilági majmokra utal, míg a Propliopithecoidea, Pliopithecoidea és Saadanius az Afro-Arábiából származó, koronás katarrhine szinapomorfiát nélkülöző törzskatarrhineákra utal.
Az élő hominoidák között fennálló ilyen adaptív hasonlóságok miatt érdekes annak vizsgálata, hogy a hominoidák más katarrinus főemlősöktől, nevezetesen a cercopithecoidáktól (óvilági majmok) való kezdeti eltérését mi jellemezte (ha jellemezte egyáltalán). A kombinált morfológiai és genetikai bizonyítékok a hominoidák és a cerkopithecoidák közötti eltérést 29,6 Ma-ra (~24-38 Ma), a hylobatidák és a főemlősök közötti eltérést pedig 18,8 Ma-ra (~16-24 Ma) teszik (Steiper és Young, 2009). A hominoidáknak tehát már az oligocénben jelen kellene lenniük, de hogyan ismerhetjük fel őket? Megtaláltuk őket? E korai fosszilis majmok azonosítását potenciálisan a következő problémák akadályozzák:
1) A hiányos fosszilis feljegyzések. Kevés olyan oligocén lelőhely van Afrikában, amely a megfelelő időszakot képviseli, és bár számos korai miocén lelőhely van, sok katarrhine taxon gyengén reprezentált, különösen a postcraniumban.
2) A Hominoidea kialakulása óta eltelt idő hossza. Egy taxon bazális tagjai kevés hasonlóságot mutathatnak a ma élő formákkal, mivel gyakran számos primitív vonást őriznek meg. Például egy nagyon korai majom még akkor is lehetett pronográd, ha az élő majmok utolsó közös őse a felegyenesedett testtartáshoz való alkalmazkodást fejlesztette ki. A törzshominoidák és a koronás hominoidák közötti potenciális szinapomorfiák tehát várhatóan kevesen vannak.
3) A ma élő vonalak hosszú evolúciós periódusai egyedileg egyediek lehettek, így a modern taxonok kevés meghatározó szinapomorfiát őriznek meg, vagy a feltételezett szinapomorfiák egy része párhuzamosság lehet.
4) Előfordulhat, hogy a morfológiai átalakulások evolúciós története nem megfelelően megoldott, ami ellentmondásos filogenetikai értelmezésekhez vezet.
Az összehasonlító vizsgálat mind a törzs-, mind a koronás katarrhinesek bevonásával enyhítheti ezeket a problémákat (Zalmout et al., 2010). Például egy késő oligocén vagy kora miocén fajnál jelenlévő, a törzskatarrhinákkal és a cercopithecoidákkal primitív módon nem közös, de a koronás hominoidáknál megfigyelhető jellegzetességek plauzibilis hominoid szinapomorfiák. Például az etmo-frontális sinus előfordul az afrikai majmoknál, és hiányzik az afro-arab törzskatarrhineusoknál, a cercopithecoidáknál, a Pongónál és a Hylobatesnél (Rossie et al., 2002; Rossie, 2008). A sinus három korai miocén kori feltételezett hominoidánál is előfordul: Proconsul, Morotopithecus és Afropithecus (ibid.). E tulajdonság elterjedése tehát úgy értelmezhető, mint egy származtatott jelleg, amely először a törzshominoidáknál fordult elő, és az ázsiai majmoknál elveszett (Begun, 1992; Moyà-Solà és Köhler, 1995; de a jelleg alternatív értelmezési lehetőségeiről lásd Rossie, 2008). Egy másik koponyatulajdonság, a szemfogak megnövekedett keresztmetszeti robusztussága és az elülső szájpadlás egyidejű kitágulása hasonló eloszlást mutat (ill, megtalálható a ma élő hominidákban (az emberben másodlagosan elveszett), valamint a Proconsulban, a Morotopithecusban és az Afropithecusban, de hiányzik a törzskatarrhineákból és a cercopithecoidákból), és szintén alátámasztja azt az értelmezést, hogy a miocén taxonok hominoidák (Begun et al., 1997; Zalmout et al., 2010). A késő oligocén Kamoyapithecus csak gnathodentális maradványokból ismert (Leakey et al., 1995), de a keresztmetszetben kitágult felső szemfogak jelenléte jelezheti, hogy ez is hominoid (Zalmout et al., 2010). Ráadásul e négy fosszilis nemzetségbe olyan fajok tartoznak, amelyek becsült testmérete meghaladja a 30 kg-ot, ezek az első catarrhine-ok, amelyek elérték ezt a méretet.
A kihalt majmok nagy méretét és arboreális niche-jét tekintve (a gibbonok valószínűleg másodlagosan törpék: Tyler, 1993), a nagy testméret szintén törzshominoid jellegzetesség lehet. A korona-hominoidáknál ezt a tulajdonságot hagyományosan a jó minőségű arboreális táplálék hominoid kiaknázásához kötik, amit a mozgásszervi innovációk tettek lehetővé, beleértve a végtagok fokozott mozgékonyságát és megragadási képességét (amelyek lehetővé teszik a súly több támaszra való elosztását), valamint az ortogradia (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin és Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Ezekre a koponya utáni átalakulásokra vonatkozó bizonyítékok a Proconsul, az Afropithecus és a Morotopithecus esetében változóan fejeződnek ki. A Proconsul több részleges csontvázról is jól ismert, és egy pronográd törzzsel rendelkező, ágak feletti négylábúként rekonstruálták, de úgy tűnik, hogy nem rendelkezett farokkal (Napier és Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). A Proconsul a rekonstrukció szerint a primitív katarrhineusokhoz és cercopithecoidákhoz képest fokozott fogási képességgel és nagyobb mozgékonysággal rendelkezett az ízületekben, például a könyökben, amit olyan jellegzetességek bizonyítanak, mint a distalis felkarcsonton található határozott zona conoidea (Rose, 1988). Az Afropithecus kevésbé jól reprezentált posztkranialisan, de ismert elemeiben hasonlít a Proconsulra (Ward, 1998). Bár a Moroto lelőhelyekről származó koponya- és koponyacsontminták közötti összefüggésről nincs egyetértés (Senut et al., 2000), a Morotopithecus koponyacsont utáni maradványai következetesen különböznek a Proconsulnak tulajdonítottaktól, ehelyett egy merev alsó háttal, mozgékony gleno-humeralis ízülettel és szándékos mászáshoz igazított combcsonttal rendelkező hominoidára utalnak, amely inkább a koronás hominoidákra emlékeztet (MacLatchy et al., 2000).
A nagyobb testméretek mellett az összes értékelhető taxonban kifejezett szexuális dimorfizmus figyelhető meg mind a testméret, mind az agyar morfológia tekintetében (Kelley, 1986; Ruff et al., 1989). Míg az erős nemi dimorfizmus primitív katarrhine jellemző, addig a látszólagos törzshominoidáknál és a modern emberszabású majmoknál továbbfejlődött (de az embernél másodlagosan csökkent).
A hominoidák fogazatában kevés, vagy egyáltalán nincs meggyőző szinapomorfia. Mivel a hominoidák megőrzik a primitív katarrinus fogmorfológiát (Kay és Ungar, 1997), míg a cercopithecoidák fogazata a bilophodontia egyértelmű szinapomorfiáján osztozik, a gyengén diagnosztikus fogazati jellegzetességek felhasználásával már régóta elismerik, hogy néhány gyengén reprezentált oligocén és miocén katarrinus hominoid lehet, amíg nem találnak teljesebb példányokat (pl. Andrews, 1978 és 1992; Stevens et al., 2013). Az ilyen oligocén és miocén katarrhinákat informálisan “fogas majmok” néven emlegetik (pl. Fleagle és Kay, 1987; Cartmill és Smith, 2009), vagy taxonómiai bizonytalanságba helyezik olyan megnevezésekkel, mint a Superfamily incertae sedis (pl. Begun, 2007).
Figyelemre méltó az az elképzelés, hogy a fent tárgyalt fosszilis taxonok egyike sem hominoid, és hogy az első felismerhető hominoid a 14 millió éves Kenyapithecus, amely a korai miocén formákhoz képest rendelkezik néhány származtatott koponyatulajdonsággal (pl. csökkent moláris cingulum, a járomív magasabb származási pozíciója) (Harrison, 2010). Ha így lenne, akkor a hominoidák becsült eredete és első megjelenése között legalább 10 Ma megmagyarázhatatlan űr tátongana – annak ellenére, hogy ezen időszak nagy részében robusztus fosszilis adatok állnak rendelkezésre -.
Bár a paleoantropológusok nem tudják biztonsággal meghatározni a “fogas majmok” vagy a Kamoyapithecus filogenetikai helyét, széleskörűen támogatják azt a hipotézist, hogy a Proconsul, Afropithecus és Morotopithecus inkább törzshominoidák, mint törzskatarrhineák. A korai hominoidák evolúciójának ablakaként ezek a taxonok a következőket sugallják:
1) A nagy testméret, az elülső fogazat megnagyobbodása és a hátsó fogazat finom változásai (pl. megnyúltabb molárisok és változások a zápfogcsúcsok elhelyezkedésében és a cingulum fejlődésében) arra utalnak, hogy alapvetően más ökológiai fülkéket foglaltak el, mint a törzskatarrhinák. A nagy szemfogak a táplálékszerzésben és a szexuális szelekcióban is szerepet játszanak, és a kifejezett dimorfizmus arra utal, hogy ez utóbbi tényező volt. A nagy méret lehetővé teszi az új táplálékhoz való hozzáférést és javítja a táplálékszerzés rugalmasságát (Wheatley, 1987), csökkenti a ragadozási kockázatot, elősegíti a túlélést, és olyan tényezőkkel párosulva, mint az élőhely stabilitása, összefüggésbe hozható a késleltetett ivaréréssel. Az Afropithecus (és valószínűleg a Proconsul) fogazati fejlődési mintázatának elemzése más emberszabásúakhoz képest késleltetett érésre utal (Kelley, 1997, 2002), ami a lassú élettartamot egy másik lehetséges törzshominoid szinapomorfiává teszi.
2) A korai miocénre a nagy testű valószínűsíthető törzshominoidák között legalább szerény mozgásszervi és táplálkozási változatosság mutatkozik, ami egy valódi adaptív radiáció kezdetét jelzi, a növekvő testméret, a növekvő mozgásszervi újdonságok és az arboreális táplálékforrások specializáltabb kiaknázása egyidejűleg fejlődik.
3) Jelenleg nem megoldható, hogy a modern majmoknál az ortogradia és az ízületi mozgékonysággal kapcsolatos anatómiai jellemzők közül melyek homológok vagy homoplasztikusak (pl. Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Ez a vita hatással van az állítólagos törzshominoidák evolúciós helyzetének értelmezésére, valamint a modern majomfélék alkalmazkodásának kialakulási mintázatának és időzítésének rekonstruálására.
Például a Morotopithecus és az élő majmok közötti koponya utáni hasonlóságok többféleképpen is értelmezhetők. Ha az élő majmok párhuzamos evolúció során szerezték meg az ortográdiát, akkor a Morotopithecus egy olyan átmenet legkorábbi ismert példáját képviseli, amely a hominoidák evolúciójában többször is bekövetkezett. Ez nagy adaptív érdekesség, ami arra utal, hogy valamilyen mögöttes fejlődési vagy genetikai hajlam (lásd alább) folyamatosan ugyanabba az irányba terelte a hominoidák evolúciós válaszát. Ez azt is jelentené, hogy bizonyos koponya utáni tulajdonságokat óvatosan kell használni a filogenetikai elemzésben (Sanders és Bodenbender, 1994). Másrészt, ha ezek a mozgásszervi hasonlóságok egy közös őstől öröklődtek, akkor a Morotopithecus közelebbi rokonságban állna a koronás hominoidákkal, mint a Proconsul (és esetleg az Afropithecus). Egy másik lehetőség, hogy a Morotopithecus egy független “kísérlet” volt, és a koronás hominoidák közös őse ennek ellenére ortográd volt.
4) Mint fentebb tárgyaltuk, a mozgásszervi specializációban mutatkozó különbségek és a független evolúció hosszú időszaka ellenére az összes fennmaradt hominoida taxon hasonló testfelépítéssel rendelkezik, ami megkülönbözteti őket más főemlősöktől (kivéve néhány atline majmot, amelyeknek konvergens módon fejlődött ki a majmokéhoz hasonló törzse és mellső végtagjainak specializációja (Erikson, 1963; Larson, 1998). A majom testfelépítésének alapvető anatómiai alapjai figyelemre méltó mozgásszervi változatosságot és sokoldalúságot eredményeztek, lehetővé téve a nagy testű főemlősök számára a hatékony arboreális (mászás és függeszkedés révén) és terrestriális (csuklós járás és kétlábúság révén) mozgást. Az ilyen mozgásszervi innováció, amely olyan tulajdonságokhoz kötődik, mint az ortogradia, valószínűleg minden élő hominoida evolúciós “sikerének” hátterében áll, és úgy tűnik, hogy bizonyos fokig már a törzshominoidáknál is jelen volt. Young és munkatársai (2010) fejlődési munkája kimutatta, hogy a négylábú majmokhoz képest a ma élő majmoknál csökkent az elülső és a hátsó végtagok közötti integráció. E genetikai kényszer lazulását úgy értelmezték, hogy a végtagok a szelekciós nyomás hatására szabadabban fejlődhettek különböző módon (ibid.). A Morotopithecus (MacLatchy et al., 2013) és a fiatalabb miocén taxonok (pl. Moya Sola és Kohler, 1996; Moya Sola et al., 2004; Nakatsukasa és Kunimatsu, 2009) ideiglenesen alátámasztja Young és munkatársai (2010) feltételezését, miszerint a végtagok “fejleszthetősége” egy ősi hominoid tulajdonság lehet, de több fosszilis anyagra és a tulajdonság hátterében álló genetika jobb megértésére van szükség.
A hominoidák eredetének és korai evolúciójának jövőbeli megismerése a lehetséges hominoid szinapomorfiák alapjainak azonosítására és megértésére irányuló további kísérletekből fog kiderülni. Ehhez viszont mind a törzs-, mind a koronás katarrhineák jobb fosszilis adataira, valamint a vizsgált közös jegyek részletesebb anatómiai, fejlődési és genetikai vizsgálatára lesz szükség. A fosszilis hominoidák paleo-környezeti kontextusban történő integrált tanulmányozása segíthet jobban megérteni azt is, hogy miért szelektálódtak a hominoidák jellemzői, akár egyszer, akár többször.