(「ヒト」/ドイツ・ネアンデル谷)

SITES
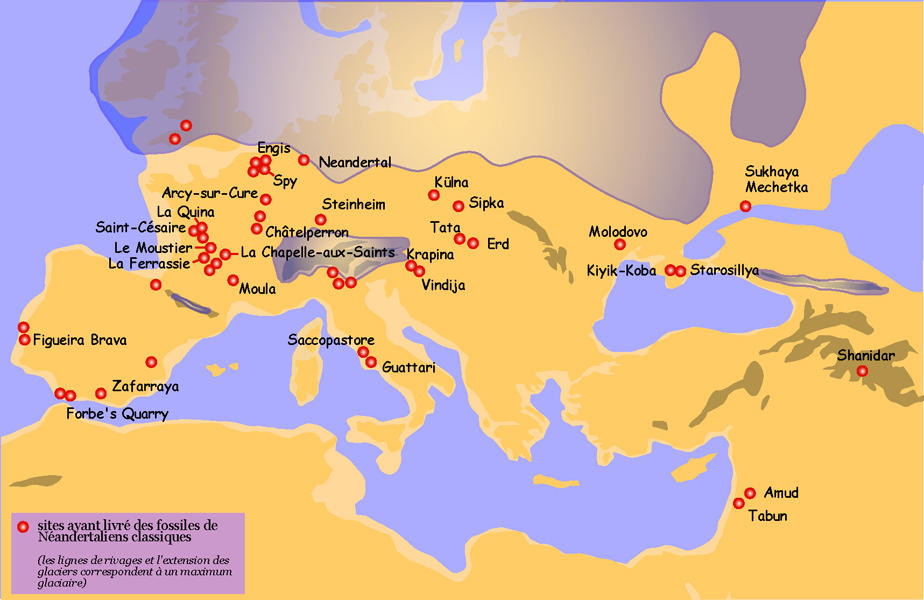
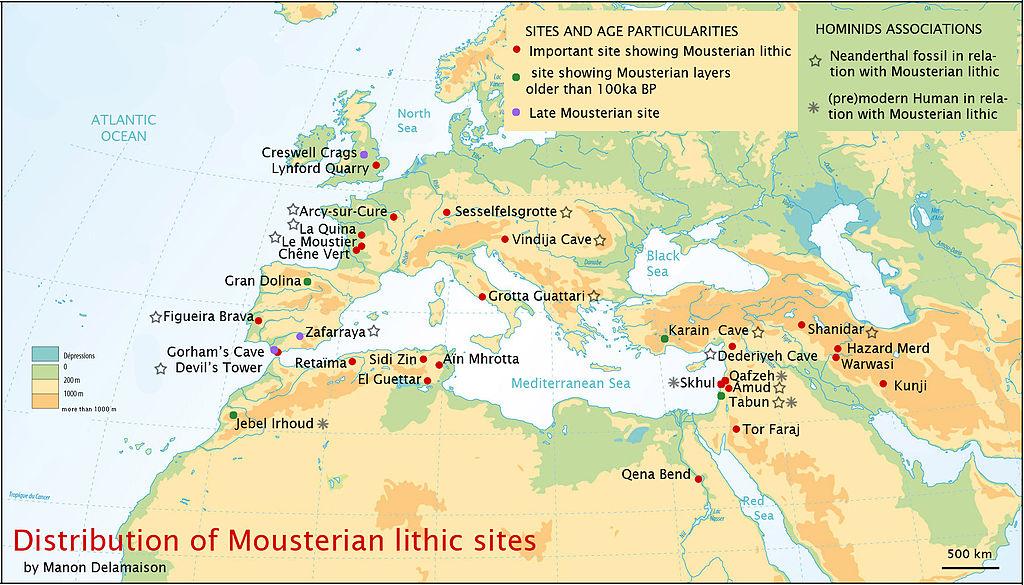
図35「ネアンデルタール遺跡」マップを参照。2
PEOPLE
挙げればきりがない(著名な研究者は本文参照)
INTRODUCTION
古生人類の中で最も知られていて最も謎めいているのが、ネアンデルタール人(Homo Neanderthalensis)であった。 ホモ・ネアンデルターレンシスはもともと我々の属と種に含まれていたが、亜種の地位、すなわちホモ・サピエンス・ネアンデルターレンシスとして区別されていた。しかし、DNA分析からの証拠が増え、2つの系統が300キロヤより前のいつか、新しいDNA証拠が正しければ、おそらく800キロヤより前に分裂したと示唆している。 しかし、DNAの証拠からは、AMHがアフリカから1回以上移住したり、中東でネアンデルタール人と同居したりして、両者が交雑していたことが示されている。 ユーラシア人とオーストラリア人は、平均して2.5%のネアンデルタール人の遺伝子を持っている。 このように、AMHの起源についてはRAOモデルが依然として支持されており、ネアンデルタールは別種と考えられているが、少なくともいくつかの集団は交雑が可能であり、その時点と地理的空間では真の生物種とは言えなかった。 ドイツ語で渓谷は「タール」といい、「h」は無声である。 一般名では “h “が省略されている資料もある。 th」を発音する人に嫌気がさした人が、それをやめさせるキャンペーンの先頭に立ったとしか思えない!
PHYLOGENY
ネアンデルタールとAMHの起源については、2つのシナリオが考えられる。 1つは化石の証拠に基づくもの、もう1つはDNAに基づくものである。 化石記録の解釈では、両種はハイデルベルゲンシスに由来し、ハイデルベルゲンシスはアフリカのエルガスターの派生型(おそらくマウリタニクス)から進化した可能性が高いとされている。 5881>
最新の遺伝的証拠は、アフリカで800キロメートル以前にAMHとネアンデルタール系統に分かれたことを支持している。 そして、新生類とデニソワ人につながる枝は、H. ハイデルベルゲンシスと考えられている。 その後、デニソワ人とネアンデルタール人は640kyaに分裂した。 しかし、局地的な集団は交配を続けた。
私たちの時代が始まって以来、ヒトの集団が交配し、遺伝的関係を維持、形成してきたことを知り、新鮮な気持ちになった。
ネアンデルタール/ヒト/デニソワの系統にかかわらず、ハイデルベルグエンシスの集団が西ヨーロッパに移動し、そこで局地的な集団が、<300kyaにネアンデルタール系統に進化しました。
更新世のヨーロッパが寒冷化するにつれて、新生児はより過酷な環境に適応した。 西ヨーロッパのネアンデルタール人は、寒さに適応した発育不全の体型をしており、東や南東のより優美な体型を保ったネアンデルタール人と区別して、「古典的」ネアンデルタール人と呼ばれる。 古典的ネアンデルタールの年代は75〜<30 kyaである。 図35.2にユーラシア大陸の新生児遺跡を示す。
DISCOVERY AND GEOGRAPHIC RANGE
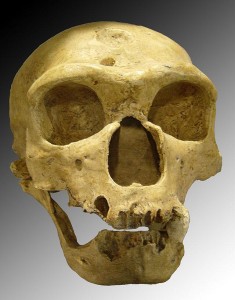
最も早く認識された発見は、ベルギーとジブラルタルで行われたものだった。 次に発見されたのがネアンデルバレーの遺跡で、これが種名の由来となった。 西ヨーロッパには化石がいたるところにあり、その大部分はフランスの水量の多い河谷に位置しています。 フランスのレエジスから半径20マイル以内に200以上の遺跡がある。 また、ドイツ、ベルギー、スペイン、ポルトガル、イタリアにもある。 有名なところでは、フランスのラ・シャペル・オ・サント、ラ・フェラッシー、サン・セゼール、ドイツのネアンデル渓谷、スペインのザファラヤ洞窟などがあります。 シャペル・オ・サン遺跡は、ネアンデルタール人が巨体で野蛮な原始人であるという神話を発展させる上で重要な役割を担っている。 1908年に発掘された約40歳の男性の遺骨(図35.3参照)は、マーセラン・ブールによって分析され、原始的で残忍、そして猫背であると特徴づけられた。 後に、この成人は関節炎を患っており、それがこの姿勢の原因であることが判明した。 ネアンデルタール人が我々人類と比較してどのように行動していたかは分からないが、彼らは前例のない文化的、技術的な複雑さを達成したのである。
西ヨーロッパが起源とされる彼らは、東は中東、遠くはウズベキスタン、北東はロシアまで、デニソワ人の地域に広がっていた。 ウズベキスタンのテシク・タシュ遺跡の9歳の少年がネアンデルタール人であるとは認めず、AMHであると主張する研究者もいます。
非古典派のネアンデルタール遺跡はクロアチア、チェコ、ハンガリー、シリア、グルジア共和国、ロシア、ウクライナ、イラク、ウズベキスタン、イスラエルで見つかっています。 有名な遺跡としては、ユーゴスラビアのKrapinaとVindija、イスラエルのKebara、Amud、Tabunの洞窟遺跡、イラクのShanidar、そして前述のウズベキスタンのTeshik Tashがある。
イスラエルの遺跡は、近くのAMH遺跡と一見同時代と思われるので何十年も興味を持たれてきた。 この2つの種の相互作用の性質については、多くの憶測がなされてきました。 一説には、氷床がヨーロッパの大部分を覆ったとき、ネアンデルタール人が他の動物とともに中東に移動したとも言われている。 AMHは120kyaには中東に到達していたが、40kyaを過ぎるまでヨーロッパに進出していなかったことから、ネアンデルタールはヨーロッパを「押さえ」、AMHの侵入を防いだとする説がある。 ネアンデルタール人は中東に移動するとき、その地域に住んでいたAMHを押し出したのかもしれない。 その後、温暖な時期になると、ネアンデルタール人がより北に移動した後、AMHが再びこの地域に移動した可能性がある。 このような交易地という考え方は、現在では、少なくともある時点ではいくつかのグループが同時期に交配していたという考え方に取って代わられている。 氷河期には集団が南下し、西ヨーロッパの集団は地中海に近づき、東のネアンデルタールは他の動物とともにイスラエルや他の温暖な地域に押し流された可能性がある。 化石の記録から、動物の群れは気候の変動に合わせて緯度を上下させたことが分かっているので、ヒトの群れも同じように移動した可能性が高い。 AMHが西ヨーロッパに移動してきた頃(約3500年前)には、ネアンデルタール人は死に始めていた。 彼らはおそらく、ますます厳しくなる気候に屈したのでしょう。 また、ある時点で進化のボトルネックを経験し、遺伝的多様性のいくつかを失い、おそらく病気に対してより脆弱になった。 中東と同様、AMHが西ヨーロッパに到着したとき、何が起こったかについては多くの憶測がある。 AMHはネアンデルタール人の遺伝子を受け継いでいた可能性が高いが、ネアンデルタール人と交配していなかった可能性もあり、西ヨーロッパの集団は中東のネアンデルタール人とは多少異なった姿をしていたことは確かであろう。 しかし、AMHが東部の地理的範囲を通過して西ヨーロッパに向かう頃には、東部のネアンデルタール人はほとんどいなくなっていたのである。 AMHは、コンテスト競争と呼ばれる直接的な競争か、スクランブル競争と呼ばれる間接的な競争、あるいは遭遇した彼らを殺してしまった可能性が指摘されている。 競争型競争とは、あるグループが他のグループの資源へのアクセスを妨げることであり、スクランブル型競争とは、あるグループが他のグループよりも資源へのアクセスに長けていることを意味する。 私はいつも、ビュッフェのテーブルを他者から守るいじめっ子と、イースターのエッグハントで卵を手に入れたり見つけたりするのが得意な子供たちのスクランブルを思い浮かべてしまうのです。 また、ネアンデルタール人は、AMHが彼らの領土を侵食したため、疎外されたことも広く受け入れられている。 クロアチアのヴィンディヤ遺跡で発見された28kyaを除いて、最近の年代はイベリア半島でのものであり、そこでネアンデルタールは後退し、絶滅したと考えられている。 この2つの種の間で何が起こったかはともかく、ネアンデルタールは退却の途にあったと思われるので、それは無意味なことなのだろう。 人間が新生物の滅亡に一役買ったと長年考えてきた結果、(少なくとも私たちが知る限り)戦争ではなく愛を育んだようだ!
PHYSICAL CHARACTERISTICS
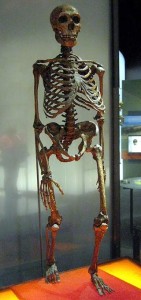
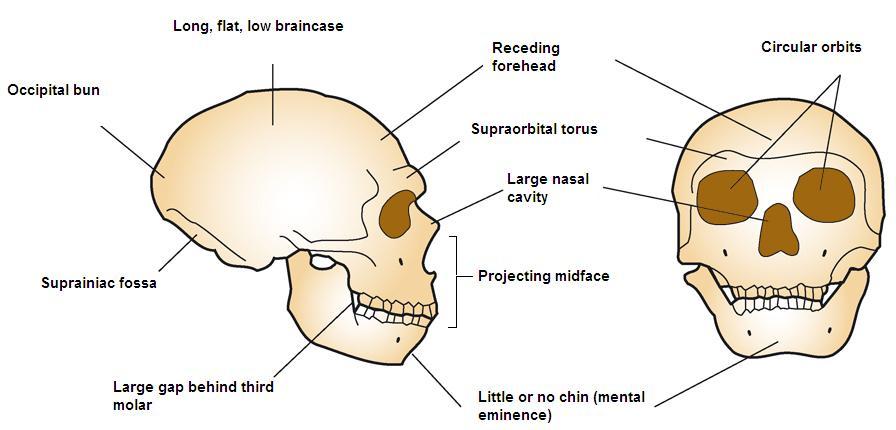
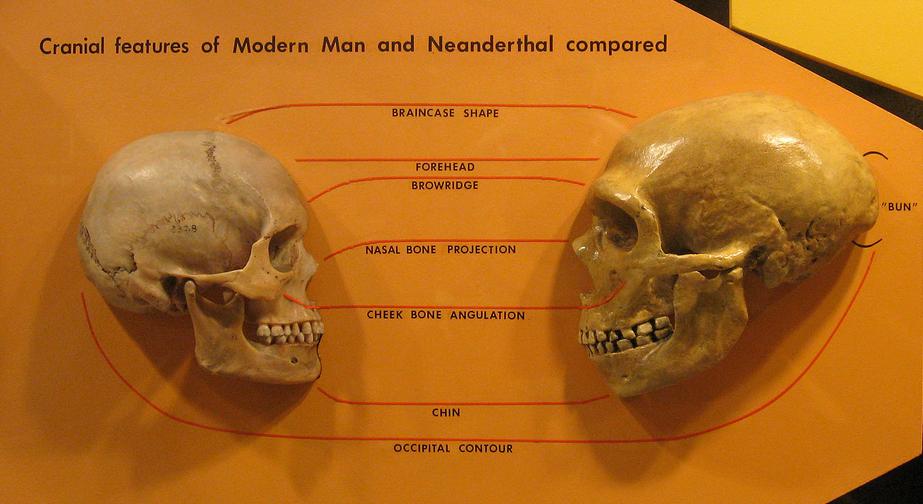
ネアンデルタールの全骨格図については図35.4を参照ください。 前述したように、西部と東部のネアンデルタールは時間の経過とともに形態的に分岐した。 そのクリンナルな変化、すなわち地理的空間における身体的特徴の段階的な変化は、時間、慢性的に寒い氷河期の条件への様々な適応と暴露、そしておそらく中東のAMHとの遺伝子流動の結果であったと考えられている。 西ヨーロッパの集団はより高緯度に住んでおり、古典的なネアンデルタールはBergmannの法則とAllenの法則に合致する寒冷適応を示した。 バーグマンの法則とは、熱損失は表面積の関数であるため、赤道から遠ざかるにつれて、熱を保存するために表面積に対して質量が増加する、というものである。 アレンの法則は、四肢の長さに関するもので、寒い環境にいる生物は付属肢が短くなる。 したがって、高温で乾燥した環境に長期にわたって適応してきた赤道アフリカの人々は、北極圏の人々の短くがっしりした体形に対して、長くしなやかな体形をしている。 ネアンデルタール人は、ずんぐりした体、短い付属肢、樽型の胸に加え、顔にも寒さに適応するための工夫をしていた。 ハイデルベルク人と同様、ネアンデルタール人も中顔面前突症、大きな鼻、副鼻腔の拡大による顔の膨張を示した。 上顎が前方に向いているため、下顎も前方に移動し、第三大臼歯の後ろに後退臼歯部と呼ばれる空間が残っていた。 鼻の内部の突起が大きく、吸入した空気を暖め、加湿するための内部表面積がさらに大きくなっている。 また、内鼻から眼窩まで独特の突起が伸びている。 寒冷適応型と東部ネアンデルタールの両方に見られるその他の頭蓋骨の特徴は、大きく、丸く、間隔の広い眼窩の上に大きく滑らかにロールした眉尾根、「後ろにそった」頬骨、いくつかの独特の内耳、後頭部には後頭部バンや上脳窩(イニオン、または外後頭隆起の上にある二つの小さなくぼみ、一般領域については図 35.5 参照)であった。 彼らの頭蓋はAMHよりも長く低い(図35.6参照)が、その絶対的な頭蓋容量は現代人をも凌駕するものであった。 バーグマンの法則によれば、脳が大きいとカロリー的にはエネルギーコストがかかるが、発熱や保温の観点からは保守的である。 新生代の脳は大きかったが、AMHの前頭葉と頭頂葉(高次の思考プロセスに関与)は新生代のそれと比べて拡張されていた。 これは氷河期のヨーロッパでAMHに優位性を与えたかもしれない。
頭蓋後のネアンデルタールは、マラソンランナー(持久力の点から)とレスラーを掛け合わせたようなものと言われている。 彼らは獲物を追い詰め、殺すために作られたのです。
ENVIRONMENT AND WAY OF LIFE
ネアンデルタールは伝統的に、厳しい気候条件に耐えてきたと描かれている。 しかし、氷河期のヨーロッパの環境に対して、どの程度の耐性があったのかについては議論がある。 Tattersall(2009)は、ネアンデルタール人は最も寒い時期にはより南方の地域に住み、気温が高くなったときにだけ高緯度に移動したとする研究の概要を示している。 彼らの形態は、伝統的な北極圏の人々のように慢性的に寒さにさらされていたことを反映しているが、そのような現代人のように優れたシェルター、仕立ての良い衣服、武器、数千年にわたる高度な文化的伝統といった近代的な技術は持ち合わせていなかったのである。 そのため、寒さに生物学的に適応する必要があった。

ネアンデルタールの集落には2種類あったことは長年知られており、あるものは一等地に住み、あるものは平原で生計を立てているかのように、しばしば偏った議論がなされたものである。 南仏の洞窟遺跡は、水利のよい河谷に、隠れ家となる洞窟、狩猟、水、道具のための石材などが豊富にあり、数千年前から人が住んでいたと考えられていた。 野外遺跡は、野外に独立した建造物を建て、家畜の群れを追って生活する、より遊牧的な集団が住んでいたようである。 洞窟遺跡と露天遺跡は、同じ民族が季節ごとに行っていた生業戦略の移り変わりを表していると考えられている。
このように、ネアンデルタール人の集団は、より温暖な地域では年間を通じて生活できたかもしれないが、それ以外の地域では彼らの祖先と同じように半遊牧民であったと思われる。 西ヨーロッパの冬の寒さには、南の洞窟に避難し、夏には移動する群れを追って北上したと思われる。 洞窟のほかに、岩壁や張り出しから外に向かって建物を建てるロックシェルターも利用された。 洞窟の中も同じように、シェルターの中にシェルターを作っていた。 地中に差し込んだ木の柱が朽ちて、腐葉土の黒っぽい輪が見えると柱状節理ができる。 この柱型の模様から、考古学的記録における住居や壁の形や大きさを復元することができる。 ネアンデルタール人は、寒冷な時期に洞窟の入り口を覆っていたことが知られており、このようなポストモールドをもとに、洞窟の入り口を覆った。 骨、柱、石を使い、皮で覆い、草で断熱していたと思われる構造物の跡が残っている。 ウクライナのモルドバ遺跡では、マンモスの骨が入った26×16´の楕円形のリングが発掘されました。
更新世において、ヨーロッパの冬は長くて寒く、夏は短くて涼しかったと言われている。 季節性があるため、植物性食品は主に暖かい時期に入手できたと思われる。 動物群や同位体分析から、トナカイとマンモスが食事の大部分を占め、ヨーロッパの新生児は高い割合で肉を食べていた。 しかし、食餌組成は地域によって異なっていた。 平地ではウマ、ウシ、ヤギが、標高の高いところではヤマヒツジやアイベックスが多く生息していた。 イラクのShanidar遺跡では、ヤギ、ヒツジ、ウシ、ブタ、カメ、クマ、シカ、キツネ、テン、スナネズミの骨が出土している。 同遺跡では、植物の消費と調理の痕跡がある。 Henry(2011)は、ネアンデルタールの歯のカルシウム沈着物(石灰質)の中にフィトリスとデンプン粒を発見した。 草のデンプンの中には、調理に特徴的な損傷が見られるものもあった。 ネアンデルタール人が火を使っていたことは、彼らの遺跡にある竈(かまど)で証明されており、植物が手に入るときにはそれを食べていたと思われるが、ようやく裏付けができたことは貴重である。

ネアンデルタールが食人を行ったかどうかについては、しばらく議論が続いていましたが、化石資料、特にフランスのムーラゲルシーの遺跡から、少なくともいくつかのグループが自分たちを食べていたことを示す説得力のある証拠が得られています。 この遺跡のネアンデルタール人の骨には、動物の骨と同じように加工された形跡がある。 骨は解体され、骨髄を取るためにハンマーで叩かれ、筋肉を切除した跡があります。
私たちにとって、共食いがどれほど忌まわしいものであるかは興味深いことです。 私たちはネアンデルタールに共感し、彼らがカニバリズムを実践していたことに失望を覚えるかもしれません。 なぜ共食いをしていたのかは不明ですが、クロアチアのクラピナなどでは、エナメル質の低形成という形で、食事によるストレスの証拠が残っています。 このように、ある集団は周期的な食糧不足に見舞われ、その結果、発育期の子供にエナメル質の異常な沈着が生じたのである。 もし人々が飢えているときに死体があれば、それを食べるというのが歴史的な証言である。 従って、ネアンデルタール人が食べるために殺すのではなく、死者を食べたとしても驚くにはあたらない。
Moula Guercy の証拠が発見されるまでは、特定の遺跡での遺体の損傷を説明するために共食いを提案されると、非常に嫌がられたものである。 ネアンデルタール人が意図的に死者を埋葬したことが、私たちの多くが親近感を抱く刺激になっているのだと思います。 人間らしいと思います。

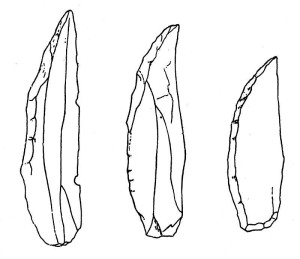
ネアンデルタール文化は、旧石器時代の中期にあたる中期旧石器時代と呼ばれる時代に属します。 新石器時代の道具は、フランスのル・ムスティエ遺跡(図35.8参照)にちなんでムステリアンインダストリー(ムステリアンツールの例は図35.9と図35.10参照)と呼ばれる。 北アフリカからはネアンデルタール人の遺跡は知られていないが、彼らの道具が発見されたことは興味深い(図35.11参照)。 ムステリアン方式は、レバロワ方式を改良したもので、得られるフレークをよりコントロールしやすくしたものである。 その結果、剥片はスクレーパーやポイントなど、さまざまな機能を持つ道具に加工された。 中には歯状、つまり鋸歯状の道具もあった。 ハイデルベルク人と同様、石器を柄や軸に取り付けて複合的な道具をつくった。 “Pointe levellois Beuzeville MHNT PRE.2009.0.203.2” by Didier Descouens is licensed under CC BY-SA 4.0.
フランスのサンセーレ遺跡から後期(35kya)道具伝統は、AMH産業の特徴を示すことから後旧石器産業にカテゴライズされる。 シャテルペロニアン伝統(図35.12参照)と呼ばれるこの伝統は、ネアンデルタールがAMHとの接触によって技術を得た、あるいは一つまたは複数の道具を見つけて独自の方法でそれを複製したことを意味し、ネアンデルタールとAMHとの直接的または間接的な接触の証拠となる可能性がある。 後者については、AMHの生産様式を学んだとする学者もいる。 フランスのアルシー・シュル・キュアの新生代遺跡から出土したいくつかの品物は宝石類と解釈されており、これもAMHにのみ起因する文化的業績である。
H. naledi と H. heidelbergensis が深い洞窟に死体を沈めたのに対し、ネアンデルタールは個々の墓に死体を埋めることが知られている最初の種であった。 遺体は屈曲した状態で発見されることが多い。 ネアンデルタール人の埋葬に関連した儀式の証拠はほとんどない。 穴を掘り、そこに遺体を折り畳み、それ故に屈曲した姿勢にし、おそらく他のものも一緒に投げ入れたと思われる。 このような遺物は、何らかの意味があると解釈されることが多いが、通常は動物の骨や壊れた道具に限られる。 しかし、ウズベキスタンのテシク・タシュ遺跡では、9歳の少年が、その体を飾っていたと思われる5本の野生のヤギの角とともに埋葬されていた。 AMHであるとの指摘もあるが、もし新生代であれば儀式的な埋葬であったようだ。
シャニダール遺跡(図35・13参照)は、私の目から見ると常に最もロマンがある遺跡であった。 定期的に陥没を経験した洞窟遺跡で、何人かの興味深い人物の遺骨が出土し、そのうちの何人かは意図的に埋葬されたものであった。 シャニダール1は成人男性である。 最終的には陥没の犠牲者となったが、彼はそれ以前の人生で1つ以上のトラウマ的な出来事を乗り越えてきた。 彼は、片方の目を失った頭部外傷のため、部分的に盲目であったと考えられている。 前腕の片方の端を失い、そのため手も失われていた。 脚は永久に不自由で、歯の一部は完全に磨り減っていた。 興味深いのは、彼はどうやって生き延びたのか、ということだ。 よく言われるのは、「仲間に助けられたから」というものだ。
Shanidar 3は、殺人の最も古い証拠かもしれません。 もう一人の成人男性である彼は、肋骨のひとつに切り傷があることから、おそらく刺されたのだろう。 骨は治った形跡があるが、この傷が原因で故意に埋められたのか、陥没して死んだのかは不明である。 もう一つ興味深い埋葬は、シャニダール4号である。 彼もまた意図的に埋葬された成人男性で、墓からは8種の野草の花粉が発見された。 種子を集めるげっ歯類の仕業とする説もあるが、ネアンデルタール人は死者を埋葬するだけでなく、遺体に花を飾っていたと考えたい人も多いだろう。
前項を書きながら、男性は女性とは異なる方法で埋葬されたのではないかと思いついた。

新人類が話すことができるかどうかという議論は、何十年にもわたって繰り広げられている。 そのため、このような「曖昧」な表現が使われるようになったのです。 私たちの喉頭は、発達の過程で下がっていく。 喉仏は、飲み物と呼吸を同時に行うために、初期には喉の高い位置にある。 赤ちゃんは、喉仏が下がるまで話すことができず、その後、喃語を話し始めます。 このように、赤ちゃんは機械的に話すことはできないが、認知的には言葉を学ぶことができる。 興味深いことに、現在では乳児に手話を教え、早くからコミュニケーションが取れるようにしている親もいます。
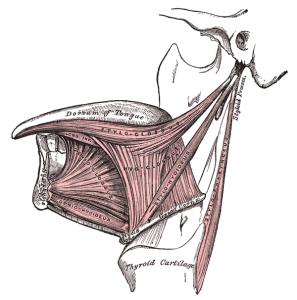
ネアンデルタールが話す能力をまだ疑っている研究者もいますが、多くの研究者は、彼らが話し言葉を持っていた可能性はあるが、我々の言葉を特徴付けるすべての音を出すことはできなかっただろうと認めています。 イスラエルのケバラ遺跡からネアンデルタール人の舌骨が発見され、その形態が我々のものと似ていることから、多くの人が彼らの言語能力を認めるようになった。 舌骨は、喉頭の靭帯や軟骨、舌の外側の筋肉(顎舌骨、舌小筋など)の重要な付着部位である。 ネアンデルタール人の音声を裏付ける最も有力な証拠は、私がこれまで述べてきたことに加え、彼らのゲノムにFOX P2遺伝子が存在していることである。 私たちもこの遺伝子を持っており、言語の習得に重要な役割を果たしています。
。