T.H. Huxley schreef dat het kijken naar andere apen is om “vage kopieën van jezelf” te zien (1863 pg. 73). De nauwe fysieke verwantschap tussen apen, of hominoïden, was al een eeuw eerder erkend door Carolus Linnaeus (1759). Levende hominoïden worden verenigd door kenmerken die verband houden met habitueel orthograaf gedrag en gedrag onder de takken: brede torso’s met ver uit elkaar staande schoudergewrichten, stijve ruggen, lange voorpoten, beweeglijke ledematen, sterk grijpend vermogen, en de afwezigheid van een staart (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Bij de mens ontbreken verschillende van deze kenmerken, zoals de relatief lange armen, als gevolg van een secundaire aanpassing aan het tweevoetig lopen. Ook schedelkenmerken verenigen hominoïden, waaronder brede voorste gehemeltes en relatief grote hersenen (Jerison, 1973; Rae, 1997). Hominoïden hebben ook een langere levensduur met vertraagde rijping (Kelley, 1997), en bezetten vergelijkbare ecologische niches – alle soorten behalve de mens bewonen tropische bossen en zijn voor hun voedsel sterk afhankelijk van rijp fruit en bladeren (Fleagle, 2013).
Hylobatiden (gibbons en siamangs) zijn kleiner dan andere levende mensapen en onderscheiden zich op andere manieren, wat hun lange periode van evolutionair isolement weerspiegelt van hominiden (mensapen en mensen), die een recentere gemeenschappelijke voorouder delen (figuur 1). Hominiden (behalve de mens) hebben een voorste verhemelte dat breder is dan dat van hylobatiden (Rae, 1997), bezitten nog kortere, stijvere onderruggen, en missen anatomische specialisaties voor ricochetale brachiation (Young, 2003).
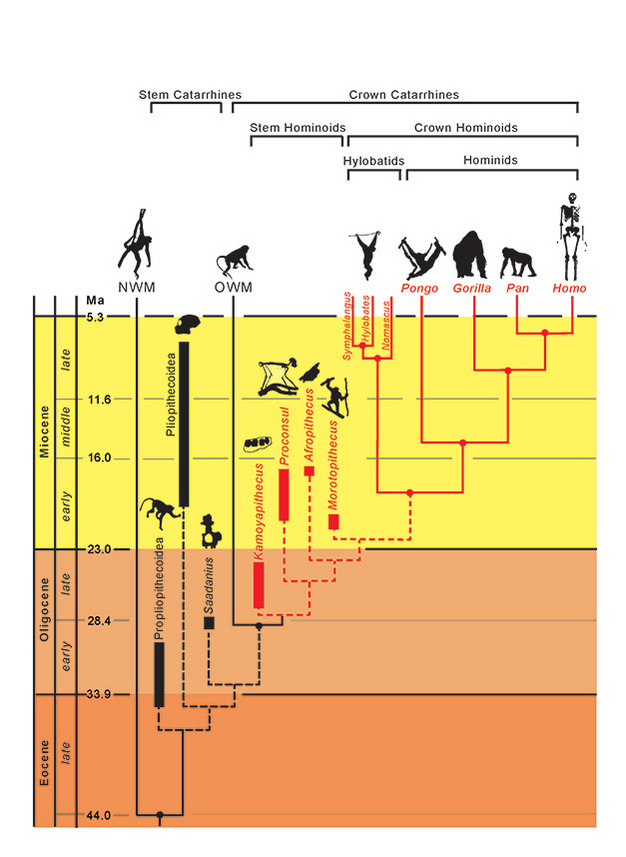
De fylogenetische relaties voor taxa worden weergegeven door ononderbroken (uitgestorven) en gestippelde (uitgestorven) lijnen. De geologische tijd wordt weergegeven op de verticale as. De timing van vertakkingen voor taxa is slechts bij benadering en afgeleid uit een combinatie van moleculaire en fossiele gegevens (Steiper et al., 2004; MacLatchy et al., 2006; Steiper en Young, 2008 en 2009; Chan et al., 2010; Stevens et al., 2013). De fylogenetische plaatsingen van de fossiele taxa zijn hypothetisch, zoals besproken in de tekst. De positie van Afropithecus en Morotopithecus zou omgekeerd kunnen zijn, of de twee zouden zustertaxa kunnen zijn. NMW verwijst naar Nieuwe Wereld apen en OWM verwijst naar Oude Wereld apen, terwijl Propliopithecoidea, Pliopithecoidea en Saadanius verwijzen naar stam catarrhines uit Afro-Arabië die geen kroon catarrhine synapomorphieën hebben.
Gezien deze adaptieve overeenkomsten tussen levende hominoïden, is het van enig belang te onderzoeken welke (indien aanwezig) de initiële divergentie van hominoïden ten opzichte van andere catarrhine primaten, met name de cercopithecoïden (apen uit de Oude Wereld), kenmerkten. Gecombineerd morfologisch en genetisch bewijs plaatst de divergentie tussen hominoïden en cercopithecoïden bij 29,6 Ma (~24-38 Ma), en de divergentie tussen hylobatiden en hominoïden bij 18,8 Ma (~16-24 Ma) (Steiper en Young, 2009). Hominoïden zouden dus al in het Oligoceen aanwezig moeten zijn, maar hoe kunnen we ze herkennen? Hebben we ze gevonden? De identificatie van deze vroege fossiele apen wordt mogelijk bemoeilijkt door de volgende problemen:
1) Een gebrekkig fossielenbestand. Er zijn weinig Oligoceen vindplaatsen in Afrika die de juiste tijdsperiode vertegenwoordigen, en hoewel er verschillende vroeg-Mioceen vindplaatsen zijn, zijn veel van hun catarrhine taxa slecht vertegenwoordigd, vooral in het postcranium.
2) De tijdsduur sedert de oorsprong van de Hominoidea. Basale leden van een taxon kunnen weinig gelijkenis vertonen met bestaande vormen, omdat zij vaak talrijke primitieve kenmerken behouden. Zo zou een zeer vroege aap pronograde kunnen zijn geweest, zelfs indien de laatste gemeenschappelijke voorouder van de levende apen aanpassingen voor een rechtopstaande houding had ontwikkeld. Potentiële synapomorfieën tussen stam- en kroonhominoïden zullen dus naar verwachting gering zijn.
3) De lange perioden van evolutie in de nog bestaande lijnen kunnen individueel uniek zijn geweest, zodat moderne taxa weinig definiërende synapomorfieën behouden, of sommige van hun veronderstelde synapomorfieën kunnen parallellen zijn.
4) De evolutionaire geschiedenis van morfologische transformatie kan niet op de juiste manier worden opgelost, wat leidt tot conflicterende fylogenetische interpretaties.
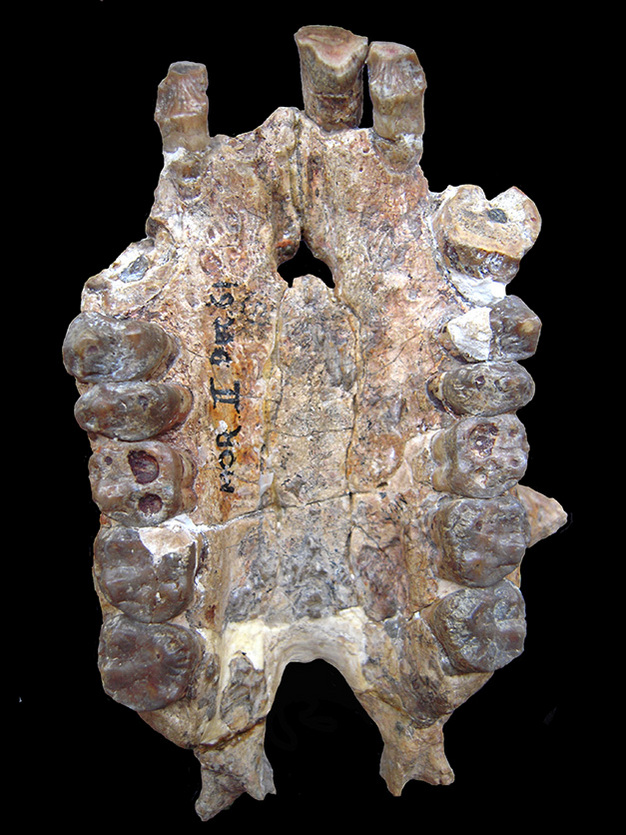
Comparatieve studie waarbij zowel stam- als krooncatarhines betrokken zijn, kan deze problemen verzachten (Zalmout et al., 2010). Bijvoorbeeld, kenmerken aanwezig in een laat Oligoceen of vroeg Mioceen soort, die niet primitief gedeeld worden met stam catarrhines en cercopithecoids, maar wel waargenomen worden bij kroon hominoids, zijn plausibele hominoide synapomorfieën. De ethmo-frontale sinus komt bijvoorbeeld voor bij Afrikaanse apen en is afwezig bij Afro-Arabische stamcatarhijnen, cercopithecoïden, Pongo, en Hylobates (Rossie et al., 2002; Rossie, 2008). De sinus komt ook voor bij drie vroeg-Mioceen vermoedelijke hominoïden: Proconsul, Morotopithecus en Afropithecus (ibid.). De verspreiding van dit kenmerk kan dus worden geïnterpreteerd als een afgeleid kenmerk dat voor het eerst voorkomt bij stamhominoïden, en verloren is gegaan bij Aziatische apen (Begun, 1992; Moyà-Solà en Köhler, 1995; maar zie Rossie, 2008, voor alternatieve manieren om dit kenmerk te interpreteren). Een ander craniaal kenmerk, de grotere robuustheid van de hoektanden en de gelijktijdige expansie van het voorste gehemelte, heeft een gelijkaardige verspreiding (d.w.z, het wordt aangetroffen bij bestaande hominiden (in tweede instantie verloren gegaan bij de mens) evenals bij Proconsul, Morotopithecus en Afropithecus, maar is afwezig bij stamcatarhines en cercopithecoïden), en ondersteunt ook de interpretatie dat de Miocene taxa hominoïden zijn (Begun et al., 1997; Zalmout et al., 2010). De laat-Oligocene Kamoyapithecus is alleen bekend van gnathodentale resten (Leakey et al., 1995), maar de aanwezigheid van doorsnede-verbrede bovenste hoektanden zou erop kunnen wijzen dat het ook een hominoïde is (Zalmout et al., 2010). Bovendien omvatten deze vier fossiele geslachten soorten met een geschatte lichaamsgrootte van meer dan 30 kg, de eerste catarhines die deze omvang bereikten.
Gezien de grote omvang en de boomachtige niche van de bestaande mensapen (gibbons zijn waarschijnlijk secundair dwerg: Tyler, 1993), kan grote lichaamsomvang ook een stam hominoide kenmerk zijn. Bij kroonhominoïden wordt deze eigenschap traditioneel in verband gebracht met de exploitatie door hominoïden van boomrijk voedsel van hoge kwaliteit, mogelijk gemaakt door innovaties op het gebied van de voortbeweging, waaronder een grotere mobiliteit van de ledematen en een groter vermogen om zich vast te grijpen (waardoor het gewicht over meerdere steunen kan worden verdeeld), en orthografie (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin en Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Aanwijzingen voor deze postcraniale transformaties komen wisselend tot uiting bij Proconsul, Afropithecus en Morotopithecus. Proconsul is bekend van verschillende partiële skeletten en is gereconstrueerd als een boven-vertakte viervoeter, met een pronograde romp, maar lijkt een staart te hebben gemist (Napier en Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). Proconsul wordt ook gereconstrueerd met een verbeterd grijpvermogen, en een grotere beweeglijkheid in gewrichten zoals de elleboog, in vergelijking met primitieve catarrhines en cercopithecoids, zoals blijkt uit kenmerken zoals een duidelijke zona conoidea in de distale humerus (Rose, 1988). Afropithecus is postcraniaal minder goed vertegenwoordigd, maar lijkt op Proconsul in bekende elementen (Ward, 1998). Hoewel er onenigheid bestaat over het verband tussen de craniale en postcraniale monsters uit de Moroto-locaties (Senut et al., 2000), verschillen de postcraniale resten van Morotopithecus consequent van die welke aan Proconsul worden toegeschreven, en suggereren in plaats daarvan een hominoide met een stijve onderrug, een beweeglijk gleno-humeraal gewricht, en een dijbeen dat is aangepast aan doelbewust klimmen, wat meer doet denken aan kroonhominoiden (MacLatchy et al., 2000).
Naast grotere lichaamsafmetingen hebben alle taxa die kunnen worden geëvalueerd een uitgesproken seksueel dimorfisme in zowel lichaamsafmetingen als in de morfologie van de hoektanden (Kelley, 1986; Ruff et al., 1989). Terwijl sterk sexueel dimorfisme een primitieve catarrhine eigenschap is, is het verder uitgewerkt in duidelijke stam hominoïden en moderne mensapen (maar secundair verminderd bij de mens).
Er zijn weinig, of geen, overtuigende synapomorfieën in het hominoïde gebit. Omdat hominoïden primitieve catarrijnse tandmorfologie behouden (Kay en Ungar, 1997), terwijl cercopithecoïde tanden de duidelijke synapomorfie van molaire bilophodontie delen, is er een geschiedenis van het gebruik van zwak diagnostische tandheelkundige kenmerken om te erkennen dat sommige slecht vertegenwoordigde Oligocene en Miocene catarrhines hominoïden zouden kunnen zijn, in afwachting van de ontdekking van meer complete exemplaren (bijv. Andrews, 1978 en 1992; Stevens et al., 2013). Dergelijke Oligoceen en Mioceen catarrhines zijn informeel aangeduid als “tandapen” (bijv. Fleagle en Kay, 1987; Cartmill en Smith, 2009), of in taxonomisch ongewisse geplaatst met een aanduiding als Superfamilie incertae sedis (bijv. Begun, 2007).
Notably, het idee is naar voren gebracht dat geen van de fossiele taxa besproken hierboven is een hominoïde, en dat de eerste herkenbare hominoïde is de 14-miljoen jaar oude Kenyapithecus, die een aantal afgeleide craniodental attributen (bijv. gereduceerde molaire cingulum, een hogere positie van oorsprong van de zygomatische boog) ten opzichte van vroege Miocene vormen (Harrison, 2010). Als dat zo is, zou er een onverklaarbare kloof van minstens 10 Ma zijn – ondanks een robuust fossielenbestand gedurende een groot deel van deze periode – tussen de geschatte oorsprong van hominoïden en hun eerste verschijning.
Hoewel paleoantropologen niet in staat zijn om de fylogenetische plaatsing van “tandapen” of Kamoyapithecus met zekerheid te bepalen, is er brede steun voor de hypothese dat Proconsul, Afropithecus en Morotopithecus stamhominoïden zijn, in plaats van stamcatarhines. Als een venster op de vroege hominoide evolutie suggereren deze taxa het volgende:
1) Grote lichaamsgrootte, vergroting van het voorste gebit, en subtiele veranderingen in het achterste gebit (b.v. meer verlengde kiezen en veranderingen in de plaatsing van de kiezen en de ontwikkeling van het cingulum) suggereren de bezetting van fundamenteel verschillende ecologische niches dan die van stamkatarrhines. Grote hoektanden spelen zowel een rol bij het verkrijgen van voedsel als bij seksuele selectie, waarbij een uitgesproken dimorfisme suggereert dat dit laatste een rol speelde. Grote afmetingen geven toegang tot nieuw voedsel en verbeteren de foerageerflexibiliteit (Wheatley, 1987), verminderen het risico van predatie, bevorderen de overlevingskansen en kunnen, in combinatie met factoren als habitatstabiliteit, verband houden met een vertraagde rijping. Analyse van het ontwikkelingspatroon van het gebit van Afropithecus (en mogelijk Proconsul) wijst op een vertraagde rijping in vergelijking met andere mensachtigen (Kelley, 1997, 2002), waardoor een langzame levensloop een andere mogelijke stamhominoïde synapomorfie is.
2) In het vroege Mioceen is er tenminste een bescheiden diversiteit in voortbeweging en dieet onder de waarschijnlijk grote stamhominoïden, wat duidt op het begin van een echte adaptieve radiatie, waarbij toenemende lichaamsgrootte, toenemende voortbewegingsvernieuwing en meer gespecialiseerde exploitatie van boomrijke voedselbronnen gelijktijdig evolueren.
3) Het is momenteel niet oplosbaar welke anatomische kenmerken geassocieerd met orthogrady en gewrichtsmobiliteit bij moderne apen homoloog of homoplastisch zijn (bijv. Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Dit debat heeft implicaties voor de interpretatie van de evolutionaire positie van vermeende stamhominoïden, en voor de reconstructie van het patroon en de timing van het ontstaan van moderne apenaanpassingen.
Bijvoorbeeld, postcraniale overeenkomsten tussen Morotopithecus en levende apen kunnen op een aantal manieren worden geïnterpreteerd. Als de levende apen hun orthogrady verwierven door parallelle evolutie, dan vertegenwoordigt Morotopithecus het vroegst bekende geval van een overgang die meerdere malen in de hominoide evolutie plaatsvond. Dit is van groot adaptief belang, omdat het suggereert dat een onderliggende ontwikkelings- of genetische neiging (zie hieronder) de evolutionaire reactie van de hominoide in dezelfde richting bleef sturen. Het zou ook betekenen dat bepaalde postcraniële kenmerken met voorzichtigheid moeten worden gebruikt in fylogenetische analyses (Sanders en Bodenbender, 1994). Aan de andere kant, als deze locomotorische overeenkomsten geërfd werden van een gemeenschappelijke voorouder, zou Morotopithecus nauwer verwant zijn aan kroon-hominoïden dan Proconsul (en mogelijk Afropithecus). Een andere mogelijkheid is dat Morotopithecus een onafhankelijk “experiment” was, en dat de gemeenschappelijke voorouder van de kroonminoïden niettemin orthograde was.
4) Zoals hierboven besproken, hebben alle overlevende hominoïde taxa, ondanks verschillen in locomotorische specialisaties en lange perioden van onafhankelijke evolutie, een vergelijkbaar lichaamsplan dat hen onderscheidt van andere primaten (met uitzondering van sommige ateline apen, die convergent een torso en voorpoot specialisaties hebben ontwikkeld die vergelijkbaar zijn met die van apen (Erikson, 1963; Larson, 1998). De anatomische basis van het apenlichaam heeft geleid tot een opmerkelijke diversiteit en veelzijdigheid van de voortbeweging, waardoor grote primaten zich doeltreffend kunnen voortbewegen in de bomen (door middel van klimmen en ophanging) en op het land (door gebruik te maken van knokkel-lopen en tweevoetig lopen). Dergelijke bewegingsinnovatie, gekoppeld aan kenmerken als orthogradie, ligt waarschijnlijk ten grondslag aan het evolutionaire “succes” van alle levende hominoïden, en lijkt tot op zekere hoogte aanwezig te zijn geweest bij de stamhominoïden. Uit ontwikkelingswerk van Young en collega’s (2010) is gebleken dat de integratie tussen de voor- en achterledematen bij bestaande mensapen is verminderd in vergelijking met viervoetige apen. Ontspanning van deze genetische beperking is geïnterpreteerd als een mogelijkheid om de ledematen vrijer te laten evolueren op verschillende manieren in antwoord op selectiedruk (ibid.). Afgeleid differentieel gebruik van ledematen in Morotopithecus (MacLatchy et al., 2013) en jongere Miocene taxa (bijv. Moya Sola en Kohler, 1996; Moya Sola et al, 2004; Nakatsukasa en Kunimatsu, 2009) ondersteunt voorlopig de veronderstelling van Young en collega’s (2010) dat “evolueerbaarheid” van ledematen een voorouderlijke hominoïde eigenschap kan zijn, maar er is meer fossiel materiaal nodig en een beter begrip van de genetica die aan deze eigenschap ten grondslag ligt.
Toekomstig inzicht in de oorsprong van hominoïden en vroege evolutie zal voortkomen uit voortdurende pogingen om de basis van mogelijke hominoïde synapomorfieën te identificeren en te begrijpen. Dit vereist op zijn beurt een beter fossielenbestand van zowel de stam- als de krooncararrhines, en een meer gedetailleerde anatomische, ontwikkelings- en genetische studie van de gemeenschappelijke kenmerken die worden onderzocht. Geïntegreerde studie van fossiele hominoïden in hun paleomilieucontext kan ons ook helpen beter te begrijpen waarom hominoïde kenmerken werden geselecteerd, hetzij eenmalig, hetzij meerdere malen.