Wprowadzenie ogólne
Większość z nas wie, że drożdże są bardzo pomocnym organizmem, szczególnie w odniesieniu do pieczenia, produkcji wina i browarnictwa. Jednakże, czym są drożdże i dlaczego są przedmiotem tak wielu badań?

Drożdże są grzybami
Drożdże są jednokomórkowymi mikroorganizmami, które są klasyfikowane, wraz z pleśniami i grzybami, jako członkowie Królestwa Grzybów. Drożdże są zróżnicowane ewolucyjnie i dlatego są klasyfikowane do dwóch odrębnych filii, Ascomycota lub grzybów workowych i Basidiomycota lub grzybów wyższych, które razem tworzą podkrólestwo Dikarya. Drożdże pączkujące, określane również jako „drożdże prawdziwe”, należą do azylu Ascomycota i rzędu Saccharomycetales. Takie klasyfikacje są oparte na cechach komórki, askospory i kolonii, a także na fizjologii komórkowej.
Drożdże są jednokomórkowe, ale mają organizację komórkową podobną do wyższych organizmów
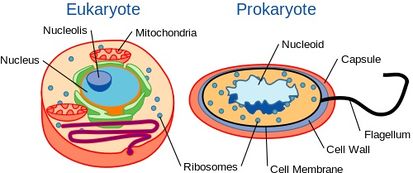
Chociaż drożdże są organizmami jednokomórkowymi, posiadają organizację komórkową podobną do organizacji organizmów wyższych, w tym ludzi. W szczególności, ich zawartość genetyczna jest zawarta w jądrze. To klasyfikuje je jako organizmy eukariotyczne, w odróżnieniu od ich jednokomórkowych odpowiedników, bakterii, które nie mają jądra i są uważane za prokariota.
Naturalne siedliska
Drożdże są szeroko rozproszone w przyrodzie z szeroką gamą siedlisk. Są one powszechnie spotykane na liściach roślin, kwiatów i owoców, jak również w glebie. Drożdże występują również na powierzchni skóry i w przewodach jelitowych zwierząt ciepłokrwistych, gdzie mogą żyć symbiotycznie lub jako pasożyty. Powszechna „infekcja drożdżowa” jest zwykle wywoływana przez Candida albicans. Oprócz bycia czynnikiem wywołującym infekcje drożdżakowe pochwy, Candida jest również przyczyną pieluszkowego zapalenia skóry oraz pleśniawek jamy ustnej i gardła.
Dlaczego warto badać drożdże?
Wyobraź sobie organizm, który szybko rośnie w kolbie i którego DNA może być łatwo manipulowane, ale również zapewnia wgląd w podstawowe procesy biologiczne człowieka, w tym choroby. Drożdże pasują do tego opisu i są obiektem badań naukowców na całym świecie, co zaowocowało ponad 50 000 opublikowanych artykułów naukowych opisujących badania nad drożdżami!

Jakie szczególne cechy drożdży sprawiają, że są one „organizmem modelowym” do badań i stanowią przedmiot tak wielu badań? Drożdże są organizmami jednokomórkowymi, co czyni je łatwymi do badania, ale posiadają organizację komórkową podobną do tej, którą można znaleźć u wyższych, wielokomórkowych organizmów, takich jak ludzie – to znaczy, posiadają jądro, a zatem są eukariotami, jak opisano powyżej. Co najważniejsze, podobieństwo w organizacji komórkowej między drożdżami i wyższymi eukariotami przekłada się na podobieństwa w ich podstawowych procesach komórkowych, więc odkrycia w drożdżach często dostarczają bezpośrednich lub pośrednich wskazówek, jak procesy biologiczne działają u ludzi.

Inną ważną cechą drożdży istotną dla ich roli jako „organizmów modelowych” jest fakt, że są one stosunkowo łatwe do pracy. Drożdże szybko się replikują i są łatwe do manipulowania genetycznego. Czas podwajania się drożdży (czas potrzebny komórce do powielenia się i podziału) wynosi około 90 minut. Dla porównania, komórki ludzkie rosnące w hodowli potrzebują około 24 godzin, aby się podwoić. Istnieją również dobrze zdefiniowane metody genetyczne dla drożdży, które pozwalają badaczom na łatwe izolowanie mutantów, krzyżowanie ich z innymi mutantami lub z innymi środowiskami genetycznymi oraz mapowanie lokalizacji genów. W rzeczywistości mapy genetyczne skonstruowane na podstawie odległości genetycznej między genami dały badaczom pierwsze spojrzenie na genom i jego organizację i były kulminacją badań genetycznych, które rozpoczęły się w pierwszej połowie XX wieku.
Przyspieszone tempo odkryć stało się możliwe po tym, jak genom drożdży piekarskich (S. cerevisiae), reprezentujący kompletny zestaw materiału genetycznego, stał się pierwszym genomem eukariotycznym, który został zsekwencjonowany w 1996 roku. Jest on mniejszy i bardziej zwarty niż genom ludzki (12 milionów par zasad i ~6 000 genów, w porównaniu z 3 miliardami par zasad i ~20-25 000 genów kodujących białka). Mimo to, porównania genomów wskazują, że ~31% genów drożdży jest bardzo podobnych do genów ludzkich, a 20% genów chorób ludzkich ma swoje odpowiedniki w drożdżach. Ponadto, komórki drożdży mogą istnieć jako haploidy (jeden zestaw chromosomów) lub diploidy (dwa zestawy chromosomów). Ponieważ haploidy mają tylko jedną kopię każdego genu oraz wydajne łamanie i ponowne łączenie nici DNA (rekombinacja), bardzo łatwo jest usunąć określony gen w haploidzie i obserwować wpływ na komórkę, czyli „fenotyp” mutanta delecyjnego. Z drugiej strony, komórki diploidalne umożliwiają badanie istotnych genów (tych, które są wymagane do wzrostu i przeżycia) poprzez usunięcie jednej kopii genu i wprowadzenie subtelnych zmian w drugiej kopii. Wreszcie, dzięki informacjom pochodzącym z sekwencji genomu, udało się stworzyć obszerny zestaw odczynników molekularnych i kolekcji obejmujących cały genom, zapewniając badaczom potężne środki do badania problemów biologicznych. Jeśli wiadomo, że gen w drożdżach jest podobny w sekwencji DNA do genu ludzkiego, badania na drożdżach mogą dostarczyć ważnych wskazówek na temat roli pokrewnego genu (genów) u ludzi. Tak więc względna prostota badania funkcji komórkowych u drożdży w połączeniu z ich znaczeniem dla organizmów wyższych czyni z nich bardzo potężny „organizm modelowy” do badań.
Życie drożdży i cykle komórkowe
Drożdże zwykle rosną bezpłciowo przez pączkowanie. Mały pąk, który stanie się komórką córką, powstaje na komórce rodzicielskiej (matce) i powiększa się wraz z dalszym wzrostem. As the córka komórka rosnąć, the matka komórka powielać i wtedy segregować swój DNA. Jądro dzieli się i wędruje do komórki córki. Kiedy pączek zawiera już jądro i osiąga określony rozmiar, oddziela się od komórki macierzystej. Serię zdarzeń, które zachodzą w komórce i prowadzą do powielania i podziału, określa się mianem cyklu komórkowego. Cykl komórkowy składa się z czterech odrębnych faz (G1, S, G2 i M) i jest regulowany podobnie jak cykl komórkowy u większych eukariontów. Tak długo jak obecne są odpowiednie składniki odżywcze, takie jak cukier, azot i fosforan, komórki drożdży będą się dzielić bezpłciowo.
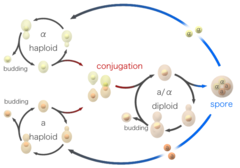
Komórki drożdży mogą również rozmnażać się płciowo. Komórki drożdży istnieją jako jeden z dwóch różnych typów kojarzenia, komórki a i komórki alfa. Kiedy komórki o przeciwnych typach kojarzenia są zmieszane razem w laboratorium lub przypadkowo stykają się w przyrodzie, mogą się łączyć (koniugować). Przed połączeniem komórki zmieniają kształt w procesie zwanym shmooing. Termin „shmoo” został ukuty na podstawie podobieństwa kształtu do fikcyjnej postaci z kreskówki o tej samej nazwie, stworzonej pod koniec lat 40-tych przez Ala Cappa, pojawiającej się najpierw w jego komiksie L’il Abner. Podczas koniugacji shmooing haploidalne komórki najpierw łączą się, a następnie ich jądra łączą się, w wyniku czego powstaje komórka diploidalna z dwiema kopiami każdego chromosomu. Po uformowaniu, komórki diploidalne mogą rozmnażać się bezpłciowo przez pączkowanie, podobnie jak haploidy. Jednakże, gdy komórki diploidalne są pozbawione substancji odżywczych, ulegają sporulacji. Podczas sporulacji komórki diploidalne przechodzą mejozę, specjalną formę podziału komórki, która redukuje liczbę chromosomów z dwóch kopii z powrotem do jednej. Po mejozie haploidalne jądra powstałe podczas mejozy są pakowane w cztery zarodniki, które zawierają zmodyfikowane ściany komórkowe, dzięki czemu powstają struktury bardzo odporne na stres środowiskowy. Zarodniki te mogą przetrwać długie okresy czasu, aż warunki staną się bardziej sprzyjające, np. w obecności lepszych składników odżywczych, kiedy to są w stanie kiełkować i rozmnażać się bezpłciowo. Te różne stany, pączkowanie, koniugacja i sporulacja razem składają się na cykl życiowy drożdży.

Wzrost i metabolizm drożdży
Kiedy komórki drożdży są hodowane w bogatych źródłach węgla, takich jak glukoza, wolą rosnąć poprzez fermentację. Podczas fermentacji glukoza jest przekształcana w dwutlenek węgla i etanol. Ogólnie rzecz biorąc, fermentacja zachodzi w nieobecności tlenu, a zatem jest z natury beztlenowa. Nawet w obecności tlenu komórki drożdży wolą rosnąć w sposób fermentacyjny, co jest określane mianem efektu Crabtree, od nazwiska biologa, który odkrył tę preferencję. Ta forma wzrostu jest wykorzystywana przy produkcji chleba, piwa, wina i innych napojów alkoholowych. Chociaż komórki drożdży pączkujących preferują wzrost w procesie fermentacji, w przypadku ograniczonej ilości składników odżywczych są one również w stanie rosnąć w procesie oddychania komórkowego. Podczas oddychania komórki przekształcają glukozę w dwutlenek węgla i wodę, zużywając w tym procesie tlen, co skutkuje produkcją znacznie większych ilości energii w postaci ATP.
Odkrycia historyczne

Drożdże są wykorzystywane jako mikroorganizm przemysłowy od tysięcy lat. Starożytni Egipcjanie wykorzystywali fermentację drożdży do wypieku chleba na zakwasie. Istnieją dowody w postaci kamieni szlifierskich, komór piekarniczych i rysunków 4000-letnich piekarni. Wykopaliska archeologiczne odkryły dowody w postaci słoików zawierających resztki wina, które mają 7000 lat.
Drożdże zostały po raz pierwszy zwizualizowane w 1680 roku przez Antoniego van Leeuwenhoeka przy użyciu wysokiej jakości soczewek. Sądził on jednak, że te kuleczki to raczej skrobiowe cząstki zboża, z których wytwarza się brzeczkę, płynny ekstrakt używany w browarnictwie, a nie komórki fermentujących drożdży. W 1789 roku Antoine Lavoisier, francuski chemik, przyczynił się do zrozumienia podstawowych reakcji chemicznych potrzebnych do produkcji alkoholu z trzciny cukrowej. Szacując proporcje materiałów wyjściowych i produktów (etanolu i dwutlenku węgla) po dodaniu pasty drożdżowej, doszedł do wniosku, że wykorzystywane są dwie ścieżki chemiczne, w których dwie trzecie cukru jest redukowane do alkoholu, a jedna trzecia do dwutlenku węgla. W tamtym czasie uważano jednak, że drożdże służą jedynie do inicjowania reakcji, a nie są wymagane w całym procesie.
W 1815 r. Joseph-Louis Gay-Lussac, francuski chemik, opracował metody utrzymywania soku winogronowego w stanie niesfermentowanym i odkrył, że wprowadzenie „fermentu” (który zawiera drożdże) było wymagane do przekształcenia niesfermentowanej brzeczki, co pokazało znaczenie drożdży dla fermentacji alkoholowej. W 1835 r. Charles Cagniard de la Tour użył mikroskopu o większej mocy, aby wykazać, że drożdże są jednokomórkowe i rozmnażają się przez pączkowanie. W latach 50. XIX wieku Ludwik Pasteur odkrył, że napoje fermentowane powstają w wyniku przekształcenia glukozy w etanol przez drożdże i zdefiniował fermentację jako „oddychanie bez powietrza”. Pod koniec XIX wieku Eduard Buchner użył bezkomórkowych ekstraktów uzyskanych przez zmielenie komórek drożdży do wykrycia zymazy, zbioru enzymów, które promują lub katalizują fermentację, za co otrzymał Nagrodę Nobla w 1907 roku.
Większość pionierskich prac nad genetyką drożdży została przeprowadzona przez Øjvinda Winge. Odkrył on, że drożdże zmieniają się pomiędzy stanami haploidalnymi i diploidalnymi oraz że drożdże są heterotaliczne, ponieważ do przekształcenia haploidów w diploidy (koniugacja) potrzebne są dwa szczepy. Wraz ze swoim współpracownikiem Otto Laustsenem opracował technikę mikromanipulacji drożdży, dzięki której można je było badać genetycznie. W tej technice, znanej jako „analiza tetrad”, cienka igła i mikroskop są używane do wyizolowania struktury znanej jako ascus, która zawiera cztery produkty zarodnikowania lub tetrad powstałe w wyniku sporulacji diploidu. Po wyizolowaniu ascusa, zarodniki w tetradzie są odrywane od siebie i pozwala się im rosnąć w kolonie do analizy genetycznej. Ta pionierska praca przyniosła mu tytuł „Ojca genetyki drożdży”. Niektóre z tych prac zostały doprecyzowane przez Carla Lindegrena, który wyjaśnił system kojarzenia w drożdżach pączkujących, wykazując istnienie komórek Mat a i Mat alfa, opracował metody masowego kojarzenia komórek tych typów i wykorzystał tę wiedzę do badania genetyki wykorzystania cukru.
Od tego czasu wielu innych badaczy przeprowadziło przełomowe badania z wykorzystaniem drożdży pączkujących. Niektórzy z tych badaczy otrzymali Nagrodę Nobla za znaczące odkrycia dokonane podczas tych badań, w tym: Dr Leland Hartwell (2001) za odkrycie genów regulujących cykl komórkowy (współzwycięzca z Paulem Nurse i Timem Huntem); Roger Kornberg (2006) za badania nad pierwszym etapem ekspresji genów, czyli sposobem, w jaki sekwencja DNA genów jest kopiowana do posłańczego RNA (mRNA); Drs. Elizabeth Blackburn, Carol Greider i Jackowi Szostakowi (2009) za odkrycie i wyjaśnienie genów i środków, za pomocą których komórki chronią końce chromosomów lub telomery przed degradacją; oraz dr Randy’emu Schekmanowi, Jamesowi Rothmanowi i Thomasowi Südhofowi (2013) za badania nad mechanizmami regulującymi ruch pęcherzykowy. Ostatnio dr Yoshinori Ohsumi otrzymał nagrodę za pracę nad autofagią, która rozpoczęła się od badań na drożdżach.
Zastosowania komercyjne

Drożdże od dawna uważane są za organizm z wyboru do produkcji napojów alkoholowych, chleba i wielu różnych produktów przemysłowych. Opiera się to na łatwości, z jaką można manipulować metabolizmem drożdży przy użyciu technik genetycznych, szybkości, z jaką można je hodować, aby uzyskać wysoką wydajność komórek (biomasę), łatwości, z jaką biomasę można oddzielić od produktów oraz wiedzy, że są one ogólnie uznane za bezpieczne (GRAS).
Drożdże pączkujące S. cerevisiae i inne gatunki drożdży są od dawna wykorzystywane do fermentacji cukrów z ryżu, pszenicy, jęczmienia i kukurydzy w celu produkcji napojów alkoholowych, takich jak piwo i wino. Istnieją dwa główne rodzaje drożdży piwowarskich: drożdże górnej fermentacji typu ale oraz drożdże dolnej fermentacji typu lager. Drożdże górnej fermentacji, takie jak S. cerevisiae wypływają na powierzchnię podczas fermentacji i są wykorzystywane do warzenia ales, porterów, stoutów i piw pszenicznych. Z kolei S. pastorianus (dawniej znane jako S. carlsbergensis) to drożdże fermentacji dolnej, używane do produkcji piwa typu lager. Drożdże lagerowe najlepiej rosną w niższych temperaturach. W związku z tym rosną wolniej, wytwarzają mniej piany na powierzchni i dlatego zwykle osiadają na dnie fermentatora. Pilznery, Märzen, Bocks i amerykańskie likiery słodowe to style piwa typu lager. W nowoczesnym browarnictwie wiele z oryginalnych szczepów górnej fermentacji zostało zmodyfikowanych, aby stać się fermentatorami dolnej fermentacji.
Drożdże produkują wino poprzez fermentację cukrów z soku winogronowego (moszczu) do etanolu. Chociaż fermentacja wina może być zainicjowana przez naturalnie występujące drożdże obecne w winnicach, wiele winiarni decyduje się na dodanie czystej kultury drożdży, aby zdominować i kontrolować fermentację. Bąbelki w szampanie i winach musujących powstają w wyniku wtórnej fermentacji, zazwyczaj w butelce, która zatrzymuje dwutlenek węgla. Dwutlenek węgla wytwarzany podczas produkcji wina jest uwalniany jako produkt uboczny. Jedna komórka drożdży może w ciągu godziny przefermentować glukozę o masie odpowiadającej jej masie. W optymalnych warunkach S. cerevisiae może wyprodukować do 18% objętości etanolu, przy czym normą jest 15 – 16%. Dwutlenek siarki obecny w komercyjnie produkowanym winie jest dodawany zaraz po zgnieceniu winogron, aby zabić naturalnie występujące bakterie, pleśń i drożdże.

Saccharomyces cerevisiae lub drożdże piekarskie są od dawna używane jako zaczyn w piekarnictwie. Drożdże piekarskie fermentują cukry obecne w cieście, wytwarzając dwutlenek węgla i etanol. Dwutlenek węgla zostaje uwięziony w małych pęcherzykach powietrza w cieście, co powoduje jego wzrost. Chleb na zakwasie jest wyjątkiem, ponieważ nie jest produkowany z użyciem drożdży piekarskich, ale z wykorzystaniem kombinacji dzikich drożdży i bakterii. Drożdże Candida milleri są używane do wzmocnienia glutenu, a bakteria wytwarzająca kwas „Lactobacillus sanfranciscensis” jest używana do fermentacji maltozy.
W uzupełnieniu do tych tradycyjnych zastosowań drożdże są również wykorzystywane do wielu innych zastosowań komercyjnych. Weganie często używają drożdży jako substytutu sera i są one często używane jako dodatek do produktów takich jak popcorn. Drożdże są wykorzystywane w przemyśle petrochemicznym, gdzie są używane do produkcji biopaliw takich jak etanol i farnezen, prekursor oleju napędowego i paliwa do silników odrzutowych. Stosuje się je również w produkcji smarów i detergentów. Drożdże są wykorzystywane w przemyśle spożywczym do produkcji dodatków do żywności, w tym barwników, przeciwutleniaczy i wzmacniaczy smaku. Drożdże są często stosowane w produkcji farmaceutyków, w tym leków przeciwpasożytniczych, związków przeciwnowotworowych, biofarmaceutyków, takich jak insulina, szczepionki i nutraceutyki. Drożdże są powszechnie stosowane w produkcji enzymów przemysłowych i chemikaliów. W dziedzinie bioremediacji środowiska szczepy były nawet wykorzystywane do usuwania metali z odpadów górniczych.
Zastosowanie w chorobach człowieka i badaniach

Dzięki wysokiemu stopniowi podobieństwa między genami drożdży i ich ludzkimi odpowiednikami oraz zachowanej podstawowej biologii komórkowej, drożdże stały się popularnym systemem modelowym do badania ludzkich genów chorobowych. Zastosowano kilka podejść, aby dowiedzieć się więcej o ludzkich genach, gdy już dojdzie do połączenia między genem ludzkim i drożdżowym. W jednym z podejść, po odkryciu ludzkiego genu związanego z chorobą, jego sekwencja jest porównywana z sekwencjami wszystkich genów w genomie drożdży w celu zidentyfikowania najbardziej podobnego genu (genów). Aby zbadać, czy geny te są funkcjonalnie powiązane, gen ludzki jest następnie ekspresjonowany w plamie drożdżowej, w której gen drożdżowy został najpierw unieczynniony przez mutację. Pozwala to badaczom określić, czy gen ludzki jest w stanie uratować żywotność, wzrost lub bardziej specyficzne defekty związane z utratą genu drożdżowego, co jest metodą określaną jako komplementacja funkcjonalna. Jeśli ścieżki i/lub procesy, w które zaangażowany jest gen drożdżowy są konserwowane, wiele można się dowiedzieć o funkcji ludzkiego genu na podstawie tego, co już wiadomo o pokrewnym genie drożdżowym. Kiedy funkcjonalna komplementarność zostanie ustanowiona, badacze mogą użyć tego systemu do dalszego scharakteryzowania funkcji produktu ludzkiego genu. Mniej ukierunkowane podejścia, które często wykorzystują techniki o wysokiej wydajności (HTP) do losowego przesiewania tysięcy ludzkich genów w jednym czasie w celu zidentyfikowania genu lub genów o aktywności komplementarnej. Takie podejścia zostały z powodzeniem wykorzystane do identyfikacji konserwatywnych regulatorów cyklu komórkowego (CDC2), genów zaangażowanych w nowotwory i genów zaangażowanych w choroby neurodegeneracyjne.
Istnieje wiele scenariuszy, w których badania mogą dostarczyć naukowcom cennych informacji na temat ścieżek komórkowych i / lub procesów, w które zaangażowany jest ludzki gen, gdy powiązany gen drożdżowy nie jest obecny. Na przykład, niektóre choroby neurodegeneracyjne, takie jak choroba Alzheimera i Parkinsona występują, gdy agregaty białkowe zwane amyloidami gromadzą się z powodu błędnego składania białek, co jest toksyczne dla neuronów. Badanie źle złożonych białek drożdżowych o podobnym potencjale tworzenia amyloidu, zwanych prionami, zapewniło naukowcom wgląd w te choroby neurodegeneracyjne. Alternatywnie, podwyższona ekspresja genu związanego z chorobą w drożdżach może skutkować fenotypem. Na przykład, przy odpowiednio wysokim poziomie ekspresji, alfa-synukleina, gen związany z chorobą Parkinsona, jest toksyczna. Taki szczep może być następnie wykorzystany do poszukiwania genów drożdży lub małych cząsteczek, które tłumią lub wzmacniają toksyczność wywołaną przez synukleinę, co często dostarcza wskazówek na temat odpowiednich szlaków komórkowych. Pacjenci ze stwardnieniem zanikowym bocznym (ALS) lub chorobą Lou Gehriga często mają mutacje w kilku białkach wiążących RNA, co powoduje, że są one podatne na tworzenie agregatów, które zakłócają metabolizm RNA. Przesiewowe badanie drożdży zostało z powodzeniem wykorzystane do zidentyfikowania wielu genów o podobnych właściwościach (tworzenie toksycznych agregatów), dostarczając naukowcom nowych genów kandydujących do badań. I odwrotnie, podczas ekspresji w drożdżach ludzkie białka wiążące RNA tworzą toksyczne agregaty, a szczep ten został wykorzystany do zidentyfikowania genu drożdżowego, który zmutowany blokuje produkcję tych agregatów.
Drożdże stają się organizmem z wyboru w badaniach mających na celu identyfikację celów farmakologicznych i sposobu działania różnych leków. Chemogenomika lub chemo-genomika odnosi się do badań, które wykorzystują połączenie chemii i genomiki do badania celów farmakologicznych i potencjalnej identyfikacji nowych leków. W badaniach chemogenomicznych stosowane są dwa główne podejścia. W pierwszym z nich konstruuje się genomową kolekcję diploidalnych szczepów, w których jedna z dwóch identycznych kopii genu jest usuwana, co obniża poziom konkretnego produktu genowego. Geny docelowe i geny zaangażowane w szlak docelowy stają się bardziej wrażliwe na dany związek i są preferencyjnie identyfikowane w tego rodzaju badaniach. W drugim podejściu nieistotne geny są systematycznie usuwane, a kolekcja badana lekiem w poszukiwaniu genów, które buforują ścieżkę docelową leku. Oczekuje się, że to podejście zidentyfikuje geny wymagane do wzrostu w obecności związku. Dodatkowe podejścia wykorzystujące ekrany z nadekspresją zostały użyte do identyfikacji genów zaangażowanych w oporność na leki, w tym potencjalny cel leku. Porównanie profilu ekspresji komórek drożdży usuniętych dla danego genu z komórkami drożdży typu dzikiego poddanych działaniu konkretnego leku może być również skutecznym sposobem identyfikacji genów, które mogą powiedzieć badaczom coś o tym, jak lek działa w komórkach.
To tylko kilka przykładów tego, jak drożdże mogą być wykorzystywane zarówno do wspomagania badań nad chorobami człowieka. Badania na drożdżach mogą pomóc badaczom dowiedzieć się więcej o biologii leżącej u podstaw choroby przy użyciu tego systemu modelowego lub pomóc im w identyfikacji celów leków lub sposobu ich działania.
Źródła
- Saccharomyces Genome Database (SGD) zapewnia kompleksowe zintegrowane informacje biologiczne dla drożdży Saccharomyces cerevisiae wraz z narzędziami do wyszukiwania i analizy tych danych.
- MIPS Comprehensive Yeast Genome Database (CYPD) przedstawia informacje o strukturze molekularnej i sieci funkcjonalnej całkowicie zsekwencjonowanego, dobrze zbadanego modelowego eukariota, drożdży Saccharomyces cerevisiae.
- Candida Genome Database (CGD), źródło danych o sekwencji genomowej oraz informacji o genach i białkach Candida albicans.
- PomBase, kompleksowa baza danych dla drożdży rozszczepialnych Schizosaccharomyces pombe, zapewniająca anotację strukturalną i funkcjonalną, kuratelę literatury i dostęp do wielkoskalowych zbiorów danych.
- Fungal genome resources at NCBI, przewodnik po zasobach genomów grzybów w National Center for Biotechnology Information (NCBI).
- Obszerna lista zasobów związanych z drożdżami na tematy od ogólnych informacji o drożdżach do kwasów nukleinowych, genomów i białek, danych o ekspresji, lokalizacji, fenotypów i innych.
Sugerowana lektura
Książki
- YeastBook. (2011) Obszerne kompendium przeglądów, które przedstawia aktualny stan wiedzy na temat biologii molekularnej, biologii komórki i genetyki drożdży Saccharomyces cerevisiae, Genetics
- From a to alpha: Yeast as a Model for Cellular Differentiation. (2007) Hiten D. Madhani, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Landmark Papers in Yeast Biology. (2006), edited by Patrick Linder, David Shore, and Michael N. Hall, Cold Spring Harbor Press, Cold Spring Harbor, New York.
- Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual. (2005) David C. Amberg, Daniel J. Burke, and Jeffrey N. Strathern. Cold Spring Laboratory Press, Cold Spring Harbor, New York.
- The Early Days of Yeast Genetics. (1993) pod redakcją Michaela N. Halla i Patricka Lindera. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Tom I: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Genome Dynamics, Protein Synthesis, and Energetics. (1991) pod redakcją James R. Broach, John R. Pringle, and Elizabeth W. Jones. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Tom II: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Gene Expression. (1992) pod redakcją Elizabeth W. Jones, John R. Pringle i James R. Broach. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Tom III: The Molecular and Cellular Biology of the Yeast Saccharomyces cerevisiae: Cykl komórkowy i biologia komórki. (1997) pod redakcją Johna R. Pringle’a, Jamesa R. Broacha i Elizabeth W. Jones. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Yeast: A Practical Approach. (1988) edited by I. Campbell and , and John H. Duffus, IRL Press, Ithaca, New York.
Journal Articles
- Duina A.A., Miller M.E., and J.B. Keeney (2014) Budding Yeast for Budding Geneticists: A Primer on the Saccharomyces cerevisiae Model System. Genetics. 197:33-48.
- Botstein D. and G.R. Fink (2011) Yeast: An Experimental Organism for 21st Century Biology. Genetics. 189:695-704.
- Cherry J.M., Hong E.L., Amundsen C., Balakrishnan R., Binkley G., Chan E.T., Christie K.R., Costanzo M.C., Dwight S.S., Engel S.R., Fisk D.G., Hirschman J.E., Hitz B.C., Karra K., Krieger C.J., Miyasato S.R., Nash R.S., Park J., Skrzypek M.S., Simison M., Weng S., and E.D. Wong (2011) Saccharomyces Genome Database: the genomics resource of budding yeast. Nucleic Acids Res. 40:D700-D705.
- Altman T.J., Boone C., Churchill G.A., Hengartner M.O., Mackay T.F., and D.L. Stemple (2011) The future of model organisms in human disease research. Nat. Rev. Genet. 18:575-582.
- Ho C.H., Piotrowski J., Dixon S.J., Baryshnikova A., Costanzo M., and C. Boone (2011) Combining functional genomics and chemical biology to identify targets of bioactive compounds. Curr Opin Chem Biol. 15:66-78.
- Smith A.M., Ammar R., Nislow C., and G. Giaever (2010) A survey of yeast genomic assays for drug and target discovery. Pharmacol Ther. 127:156-164.
- Doliński K., and D. Botstein (2007) Orthology and functional conservation in eukaryotes. Annu. Rev. Genet. 41:465-507.
- Barnett J.A. (2007) A history of research on yeasts 10: foundations of yeast genetics. Yeast 24:799-845.
- Spradling A., Ganetsky B., Hieter P., Johnston M., Olson M., Orr-Weaver T., Rossant J., Sanchez A., and R. Waterston (2006) New roles for model genetic organisms in understanding and treating human disease: report from the 2006 Genetics Society of America meeting. Genetics 172:2025-2032.
- Sherman F. (2002) Getting started with yeast Methods Enzymol. 350:3-41.