T.H. Huxley skrev att när man ser på andra apor ser man ”suddiga kopior av sig själv” (1863 s. 73). Det nära fysiska släktskapet mellan apor, eller hominoider, hade redan erkänts ett århundrade tidigare av Carolus Linné (1759). Levande hominoider förenas av egenskaper som har att göra med vanemässig ortograd och beteenden under grenarna: bred torso med brett utspridda axelleder, styva ryggar, långa framben, rörliga lemmar, stark greppförmåga och avsaknad av svans (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Människor saknar flera av dessa egenskaper, t.ex. relativt långa armar, som ett resultat av en sekundär anpassning till bipedalism. Kraniala attribut förenar också hominoider, inklusive breda främre gommen och relativt stora hjärnor (Jerison, 1973; Rae, 1997). Hominoider har också en längre livslängd med fördröjd mognad (Kelley, 1997) och upptar liknande ekologiska nischer – alla arter utom människan bebor tropiska skogar och förlitar sig i hög grad på mogna frukter och blad som föda (Fleagle, 2013).
Hylobatider (gibboner och siamanger) är mindre än andra levande apor och skiljer sig på andra sätt, vilket återspeglar deras långa period av evolutionär isolering från hominider (människoapor och människor), som delar en nyare gemensam förfader (figur 1). Hominiderna (utom människan) har främre gommen som är bredare än hos hylobatiderna (Rae, 1997), har ännu kortare och styvare nedre ryggar och saknar anatomiska specialiseringar för ricochetal brachiation (Young, 2003).
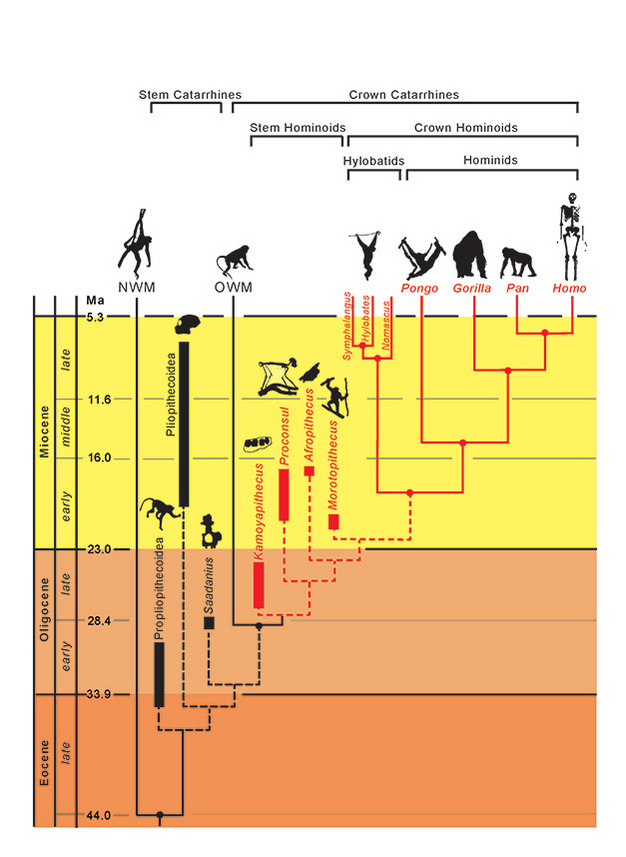
De fylogenetiska relationerna för taxa representeras av heldragna (utdöda) och streckade (utdöda) linjer. Geologisk tid representeras på den vertikala axeln. Tidpunkten för förgreningshändelser för taxa är endast ungefärlig och härledd från en kombination av molekylära och fossila data (Steiper et al., 2004; MacLatchy et al., 2006; Steiper och Young, 2008 och 2009; Chan et al., 2010; Stevens et al., 2013). Den fylogenetiska placeringen av de fossila taxa är hypotetisk, vilket diskuteras i texten. Placeringen av Afropithecus och Morotopithecus kan vara omvänd, eller så kan de två vara systertaxa. NMW avser apor från Nya världen och OWM avser apor från Gamla världen, medan Propliopithecoidea, Pliopithecoidea och Saadanius avser stamkatarrhiner från Afro-Arabien som saknar synapomorfier av kronkatarrhiner.
Med tanke på dessa adaptiva likheter bland de levande hominoiderna är det av visst intresse att undersöka vilka (om några) som kännetecknade den inledande divergensen mellan hominoiderna och andra katarrafiska primater, särskilt cercopithecoiderna (aporna från Gamla världen). Kombinerade morfologiska och genetiska bevis placerar hominoid-cercopithecoid-divergensen vid 29,6 Ma (~24-38 Ma), och hylobatid-hominid-divergensen vid 18,8 Ma (~16-24 Ma) (Steiper och Young, 2009). Hominoider bör därför finnas vid Oligocen, men hur kan vi känna igen dem? Har vi hittat dem? Att identifiera dessa tidiga fossila apor hindras potentiellt av följande problem:
1) Ett bristfälligt fossilregister. Det finns få oligocena platser i Afrika som representerar den lämpliga tidsperioden, och även om det finns flera tidiga miocena platser är många av deras katarrhina taxa dåligt representerade, särskilt i postcranium.
2) Tidsperioden sedan Hominoidea uppstod. Basala medlemmar av ett taxon kan ha liten likhet med existerande former eftersom de ofta behåller många primitiva drag. En mycket tidig apa kan till exempel ha varit pronograd även om den sista gemensamma förfadern till levande apor hade utvecklat anpassningar för upprätt hållning. Potentiella synapomorfier mellan stam- och kronhominoider förväntas därför vara få.
3) De långa utvecklingsperioderna i de bevarade släktena kan ha varit individuellt unika så att moderna taxa behåller få definierande synapomorfier, eller så kan en del av deras förmodade synapomorfier vara parallellismer.
4) Den morfologiska omvandlingens evolutionära historia kanske inte löses på lämpligt sätt, vilket leder till motstridiga fylogenetiska tolkningar.
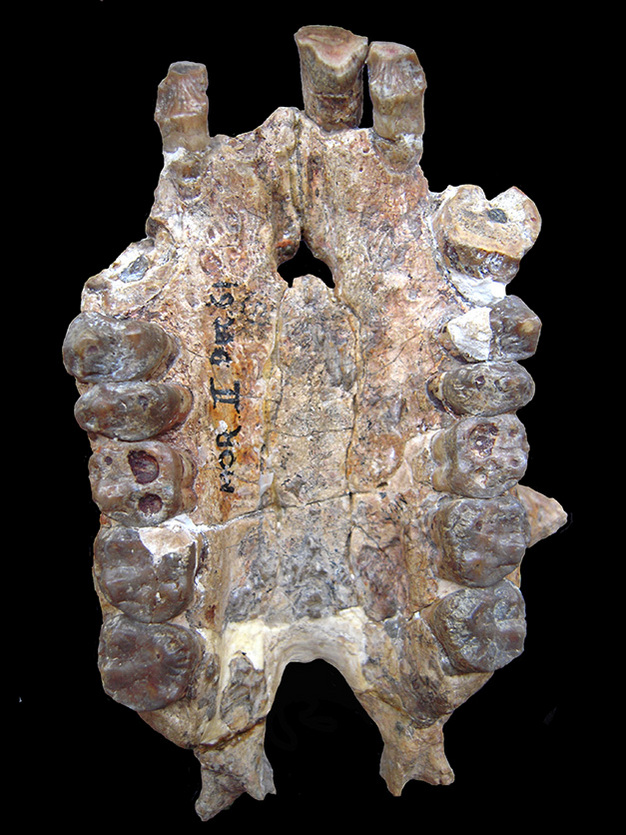
Komparativa studier som både omfattar stam- och kronkatarrhiner kan mildra dessa problem (Zalmout et al., 2010). Exempelvis är egenskaper som finns hos en sen oligocen eller tidig miocen art, som inte delas primitivt med stamkatarrhiner och cercopithecoider, men som observeras hos kronhominoider, plausibla hominoida synapomorfier. Till exempel förekommer den ethmo-frontala sinus hos afrikanska apor och saknas hos afroarabiska stamkatarrhiner, cercopithecoider, Pongo och Hylobates (Rossie et al., 2002; Rossie, 2008). Sinus förekommer också hos tre putativa hominoider från tidig miocen: Proconsul, Morotopithecus och Afropithecus (ibid.). Utbredningen av detta drag kan således tolkas som en härledd karaktär som först uppträder hos stamhominoider och försvinner hos asiatiska apor (Begun, 1992; Moyà-Solà och Köhler, 1995; men se Rossie, 2008, för alternativa sätt att tolka denna karaktär). Ett annat kraniekt drag, ökad tvärsnittsrobusthet hos hörntänderna och samtidig expansion av den främre gommen, har en liknande fördelning (dvs, det finns hos bevarade hominider (förlorat sekundärt hos människor) samt hos Proconsul, Morotopithecus och Afropithecus, men saknas hos stamkatarrhiner och cercopithecoider), och stödjer också tolkningen att de miocena taxa är hominoider (Begun et al., 1997; Zalmout et al., 2010). Den sena oligocena Kamoyapithecus är endast känd från gnathodentala lämningar (Leakey et al., 1995), men närvaron av tvärsnittsutvidgade övre hörntänder skulle kunna signalera att den också är en hominoid (Zalmout et al., 2010). Dessutom innehåller dessa fyra fossila släkten arter med uppskattade kroppsstorlekar på över 30 kg, de första katarrhiner som uppnådde denna storlek.
Med tanke på de existerande apornas stora storlek och arboristiska nisch (gibboner är troligen sekundärt dvärglika: Tyler, 1993) kan stor kroppsstorlek också vara ett stamhominoidkännetecken. Hos kronhominoider är detta drag traditionellt knutet till hominoidernas exploatering av högkvalitativ mat i träd, vilket möjliggjordes av lokomotoriska innovationer, inklusive förbättrad rörlighet och greppförmåga hos lemmarna (vilket gör det möjligt att fördela vikten på flera olika stöd) och ortograd (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin och Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
Bevisen för dessa postkraniella omvandlingar är varierande uttryckta hos Proconsul, Afropithecus och Morotopithecus. Proconsul är välkänd från flera partiella skelett och har rekonstruerats som en ovanliggande fyrbenta, med pronograd bål, men tycks ha saknat svans (Napier och Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). Proconsul är också rekonstruerad för att ha förbättrad greppförmåga och större rörlighet i leder som armbågen, i förhållande till primitiva catarrhiner och cercopithecoider, vilket framgår av egenskaper som en distinkt zona conoidea i det distala överarmsbenet (Rose, 1988). Afropithecus är mindre väl representerad postkraniellt, men liknar Proconsul i kända element (Ward, 1998). Även om sambandet mellan kranie- och postkraniellproverna från Moroto-lokalerna är föremål för oenighet (Senut et al., 2000), skiljer sig de postkraniella kvarlevorna av Morotopithecus konsekvent från dem som tillskrivs Proconsul, vilket i stället antyder en hominoid med styv nedre del av ryggen, rörlig gleno-humeral led och ett lårben som är anpassat för avsiktlig klättring, vilket påminner mer om kronhominoider (MacLatchy et al, 2000).
Utöver större kroppsstorlekar har alla taxa som kan utvärderas en uttalad sexuell dimorfism i både kroppsstorlek och hundmorfologi (Kelley, 1986; Ruff et al., 1989). Medan stark sexuell dimorfism är ett primitivt catarrhine-egenskapp, är den ytterligare utarbetad hos uppenbara stamhominoider och moderna människoapor (men sekundärt reducerad hos människor).
Det finns få, om ens några, övertygande synapomorfier i hominoidernas tandsystem. Eftersom hominoider behåller en primitiv katarrhintandsmorfologi (Kay och Ungar, 1997), medan cercopithecoid-tänderna delar den tydliga synapomorfin molar bilophodonty, finns det en historia av att använda svagt diagnostiska dentala kännetecken för att erkänna att vissa dåligt representerade oligocena och miocena katarrhiner skulle kunna vara hominoider, i väntan på att mer kompletta exemplar ska upptäckas (t.ex. Andrews, 1978 och 1992; Stevens m.fl., 2013). Sådana oligocena och miocena katarrhiner har informellt kallats ”tandapor” (t.ex. Fleagle och Kay, 1987; Cartmill och Smith, 2009), eller placerats i taxonomiskt limbo med hjälp av en beteckning som Superfamily incertae sedis (t.ex. Begun, 2007).
Noterligen har idén framförts att ingen av de fossila taxa som diskuterats ovan är en hominoid, och att den första igenkännbara hominoiden är den 14 miljoner år gamla Kenyapithecus, som har vissa avledda kraniodentala attribut (t.ex. reducerad molar cingulum, en högre position för ursprunget av zygomatiska bågen) i förhållande till tidiga miocena former (Harrison, 2010). Om så är fallet skulle det finnas en oförklarlig lucka på minst 10 Ma – trots ett robust fossilregister under en stor del av denna tidsperiod – mellan hominoidernas uppskattade ursprung och deras första framträdande.
Och även om paleoantropologer inte kan fastställa den fylogenetiska placeringen av ”tandapor” eller Kamoyapithecus med säkerhet, finns det ett brett stöd för hypotesen att Proconsul, Afropithecus och Morotopithecus är stamhominoider, snarare än stamkatarrhiner. Som ett fönster i den tidiga hominoida evolutionen föreslår dessa taxa följande:
1) Stor kroppsstorlek, utvidgning av det främre bettet och subtila förändringar i det bakre bettet (t.ex. mer långsträckta kindtänder och förändringar i placeringen av kindtandshuvudet och utvecklingen av cingulum) tyder på att de ockuperar fundamentalt annorlunda ekologiska nischer än stamkatarrhiner. Stora hörntänder fungerar både för att skaffa föda och för sexuellt urval, med uttalad dimorfism som tyder på att det sistnämnda var en faktor. Stor storlek ger tillgång till ny föda och ökar flexibiliteten vid födosök (Wheatley, 1987), minskar rovdjursriskerna, främjar överlevnaden och kan, i kombination med faktorer som stabilitet i livsmiljön, kopplas till fördröjd mognad. Analys av tandutvecklingsmönstret hos Afropithecus (och möjligen Proconsul) tyder på fördröjd mognad i förhållande till andra antropoider (Kelley, 1997, 2002), vilket gör långsam livshistoria till en annan möjlig stamhominoid synapomorfi.
2) Vid den tidiga miocen finns det åtminstone en blygsam lokomotorisk och dietär mångfald bland de storkroppiga sannolika stamhominoiderna, vilket tyder på början av en sann adaptiv strålning, med ökande kroppsstorlek, ökande lokomotorisk nyhet och mer specialiserat utnyttjande av trädlevande födokällor som utvecklas samtidigt.
3) Det går för närvarande inte att lösa vilka anatomiska egenskaper som är förknippade med ortograd och ledrörlighet hos moderna apor som är homologa eller homoplastiska (t.ex. Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Denna debatt har konsekvenser för tolkningen av den evolutionära positionen hos påstådda stamhominoider och för rekonstruktionen av mönstret och tidpunkten för uppkomsten av moderna apanpassningar.
Till exempel kan postkraniella likheter mellan Morotopithecus och levande apor tolkas på ett antal olika sätt. Om de levande aporna fick sin ortograd genom parallell utveckling så representerar Morotopithecus det tidigaste kända exemplet på en övergång som inträffade flera gånger under hominoidernas utveckling. Detta är av stort adaptivt intresse, eftersom det tyder på att någon underliggande utvecklingsmässig eller genetisk benägenhet (se nedan) fortsatte att styra den hominoida evolutionära responsen i samma riktning. Det skulle också innebära att vissa postkraniella egenskaper måste användas med försiktighet i fylogenetiska analyser (Sanders och Bodenbender, 1994). Å andra sidan, om dessa lokomotoriska likheter ärvdes från en gemensam förfader skulle Morotopithecus vara närmare besläktad med kronhominoider än Proconsul (och möjligen Afropithecus). En annan möjlighet är att Morotopithecus var ett oberoende ”experiment” och att kronhominoidernas gemensamma förfader ändå var ortograd.
4) Som diskuterats ovan har alla överlevande hominoida taxa, trots skillnader i lokomotoriska specialiseringar och långa perioder av oberoende evolution, en liknande kroppsplan som skiljer dem från andra primater (med undantag för vissa atelinjeapor, som konvergent har utvecklat överkroppar och frambensspecialiseringar som liknar dem hos aporna (Erikson, 1963; Larson, 1998). De grundläggande anatomiska förutsättningarna för apornas kroppsplan har lett till en anmärkningsvärd mångfald och mångsidighet när det gäller lokomotorik, vilket gör det möjligt för stora primater att förflytta sig effektivt i träd (genom klättring och upphängning) och på jorden (med hjälp av knäböjning och bipedalism). Sådana lokomotiva innovationer, som är knutna till egenskaper som ortograd, ligger sannolikt till grund för den evolutionära ”framgången” för alla levande hominoider, och verkar ha funnits i viss utsträckning hos stamhominoiderna. Utvecklingsarbete av Young och kollegor (2010) har visat att de existerande aporna har minskad integration mellan fram- och bakbenet jämfört med fyrbenta apor. En lättnad av denna genetiska begränsning har tolkats som att den har gjort det möjligt för lemmarna att utvecklas friare på olika sätt som svar på urvalstryck (ibid.). Uttryckt differentiell användning av lemmar hos Morotopithecus (MacLatchy et al., 2013) och yngre miocena taxa (t.ex. Moya Sola och Kohler, 1996; Moya Sola et al, 2004; Nakatsukasa och Kunimatsu, 2009) stödjer preliminärt Young och kollegors (2010) antagande att limb ”evolvability” kan vara en förfädrad hominoid egenskap, men mer fossilt material och en bättre förståelse av den genetik som ligger till grund för denna egenskap, behövs.
Framtida insikter om hominoidernas ursprung och tidiga evolution kommer att framträda genom fortsatta försök att identifiera och förstå grunden för möjliga hominoida synapomorfier. Detta kommer i sin tur att kräva ett förbättrat fossilregister för både stam- och kronkatarrhiner och mer detaljerade anatomiska, utvecklingsmässiga och genetiska studier av de gemensamma karaktärerna som undersöks. Integrerade studier av fossila hominoider i deras paleomiljösammanhang kan också hjälpa oss att bättre förstå varför hominoida egenskaper valdes ut, antingen en gång eller flera gånger.