Hormonien vapautuminenEdit
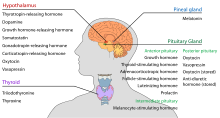
Hypotalamuksella on keskeinen neuroendokriininen funktio, joka ilmenee ennen kaikkea siten, että se kontrolloi aivolisäkkeen etulohkoa, joka vuorostaan säätelee erilaisia sisäeritysrauhasia ja elimiä. Vapauttavia hormoneja (joita kutsutaan myös vapauttaviksi tekijöiksi) tuotetaan hypotalamuksen ytimissä, minkä jälkeen ne kulkeutuvat aksoneja pitkin joko keskiaivolisäkkeeseen tai aivolisäkkeen takaosaan, jossa ne varastoituvat ja vapautuvat tarpeen mukaan.
Aivolisäkkeen etuosa
Hypotalamus-aivolisäkkeen akselissa vapautuvat vapauttajahormonit, joita kutsutaan myös hypofysiotrooppisiksi tai hypotalamushormoneiksi, vapautuvat median eminenssistä, joka on hypotalamuksen jatke, hypofyysiseen portaalijärjestelmään, joka kuljettaa ne aivolisäkkeen etuosaan, jossa ne käyttävät säätelytehtäviään aivolisäkkeen adenohypofyysihormonien eritykseen. Näitä hypofysiotrooppisia hormoneja stimuloivat hypotalamuksen periventrikulaarisella alueella sijaitsevat parvocellulaariset neurosekretoriset solut. Vapaututtuaan kolmannen kammion kapillaareihin hypofysiotrooppiset hormonit kulkeutuvat niin sanotun hypotalamus-aivolisäkkeen portaalikierron kautta. Saavuttuaan määränpäähänsä aivolisäkkeen etuosaan nämä hormonit sitoutuvat tiettyihin reseptoreihin, jotka sijaitsevat aivolisäkkeen solujen pinnalla. Riippuen siitä, mitkä solut aktivoituvat tämän sitoutumisen kautta, aivolisäke joko alkaa erittää tai lopettaa hormonien erittymisen muuhun verenkiertoon.
| Sekretoituva hormoni | Lyhenne | Tuotettu | Vaikutus |
|---|---|---|---|
| Tirotropiinia vapauttava hormoni (Prolaktiinia vapauttava hormoni) |
TRH, TRF, tai PRH | Paraventrikulaarisen ytimen parvocellulaariset neurosekretoriset solut | Stimuloivat kilpirauhasta stimuloivan hormonin (TSH) vapautumista aivolisäkkeen etupuolelta (ensisijaisesti) Stimuloivat prolaktiinin vapautumista aivolisäkkeen etupuolelta |
| Corticotropin-vapauttava hormoni | CRH tai CRF | Paraventrikulaarisen ytimen paraventrikulaariset neurosekretoriset solut | Stimuloivat adrenokortikotrooppisen hormonin (ACTH) vapautumista aivolisäkkeen etuosasta |
| Dopamiini (Prolaktiinininhiboiva hormoni) |
DA tai PIH | Dopamiinineuronit kaarevassa ytimessä | Estäisivät prolaktiinin vapautumista aivolisäkkeen etupuolelta |
| Kasvua-hormonia vapauttava hormoni | GHRH | Arcuate nucleuksen neuroendokriiniset neuronit | Stimuloivat kasvun-hormonin (GH) vapautumista aivolisäkkeen etuosasta |
| Gonadotropiinia vapauttava hormoni | GnRH tai LHRH | Preoptisen alueen neuroendokriiniset solut | Stimuloivat follikkelia-stimuloivan hormonin (FSH) vapautumista aivolisäkkeen etupuolelta Stimuloi luteinisoivan hormonin (LH) vapautumista aivolisäkkeen etupuolelta |
| Somatostatiini (kasvuhormonia estävä hormoni) |
SS, GHIH, tai SRIF | Periventrikulaarisen ytimen neuroendokriiniset solut | Estäisivät kasvuhormonin (GH) vapautumista aivolisäkkeen etuosasta Estäisivät (kohtalaisesti) kilpirauhasta stimuloivan hormonin (TSH) vapautumista aivolisäkkeen etuosasta |
Muita mediaanieminenssistä erittyviä hormoneja ovat mm. vasopressiini, oksitosiini ja neurotensiini.
Aivolisäkkeen takaosa
Hypotalamus-hypofyysi-akselissa neurohypofyysihormoneja vapautuu verenkiertoon aivolisäkkeen takaosasta, joka on itse asiassa hypotalamuksen jatke.
| Sekretoituva hormoni | Lyhenne | Tuotetaan | Vaikutus |
|---|---|---|---|
| Oksitosiini | OXY tai OXT | Paraventrikulaarisen ytimen ja supraoptisen ytimen magnosellulaariset hermosolut. Ydin | Kohdun supistuminen Imetys (laskeutumisrefleksi) |
| Vasopressiini (antidiureettinen hormoni) |
ADH tai AVP | Paraventrikulaarisen ytimen magnosellulaariset ja parvosellulaariset hermosolut, supraoptisen ytimen magnosellulaariset solut | Munuaisen distaalisen tubuluksen ja keräyskanavan solujen vedenläpäisevyyden lisääntyminen ja siten veden takaisinimeytymisen ja väkevän virtsan erittymisen mahdollistaminen |
Tiedetään myös, että hypotalamus-aivolisäke-lisämunuaiskuori-akselin (HPA-akselin) hormoneilla on yhteys tiettyihin ihosairauksiin ja ihon homeostaasiin. On näyttöä siitä, että HPA-hormonien hyperaktiivisuus yhdistetään stressiin liittyviin ihosairauksiin ja ihokasvaimiin.
StimulaatioEdit
Hypotalamus koordinoi monia hormonaalisia ja käyttäytymiseen liittyviä vuorokausirytmejä, monimutkaisia neuroendokriinisten ulostulojen malleja, monimutkaisia homeostaattisia mekanismeja ja tärkeitä käyttäytymismalleja. Hypotalamuksen on siis reagoitava moniin erilaisiin signaaleihin, joista osa syntyy ulkoisesti ja osa sisäisesti. Joko talamuksesta tai aivokuorelta tuleva delta-aaltosignaalinanto vaikuttaa vapauttajahormonien eritykseen; GHRH ja prolaktiini stimuloituvat, kun taas TRH estyy.
Hypotalamus reagoi:
- valoon: Päivänpituus ja valojakso vuorokausi- ja vuodenaikarytmien säätelyyn
- Hajuärsykkeet, mukaan lukien feromonit
- Steroidit, mukaan lukien sukurauhassteroidit ja kortikosteroidit
- Neuraalisesti välittyvä informaatio, joka on peräisin erityisesti sydämestä, suolistohermostosta (ruoansulatuskanavan hermojärjestelmästä) ja lisääntymiskanavasta.
- Autonomiset tulot
- Verestä peräisin olevat ärsykkeet, mukaan lukien leptiini, greliini, angiotensiini, insuliini, aivolisäkkeen hormonit, sytokiinit, plasman glukoosipitoisuudet ja osmolaarisuus jne.
- Stressi
- Haittavat mikro-organismeja nostamalla kehon lämpötilaa, nollaamalla kehon termostaattia ylöspäin.
HajuärsykkeetTiedosto
Hajuärsykkeet ovat tärkeitä sukupuoliselle lisääntymiselle ja neuroendokriiniselle toiminnalle monissa lajeissa. Jos esimerkiksi tiineenä oleva hiiri altistetaan ”vieraan” uroksen virtsalle kriittisenä aikana yhdynnän jälkeen, raskaus epäonnistuu (Brucen vaikutus). Naarashiiri muodostaa yhdynnän aikana kumppanistaan tarkan ”hajumuistin”, joka säilyy useita päiviä. Feromonivihjeet auttavat monilla lajeilla tiineyden synkronoinnissa; naisilla myös synkronoidut kuukautiset voivat johtua feromonivihjeistä, joskin feromonien rooli ihmisillä on kiistanalainen.
Veren välityksellä tulevat ärsykkeetEdit
Peptidihormoneilla on tärkeitä vaikutuksia hypotalamukseen, ja sitä varten niiden on läpäistävä veri-aivoeste. Hypotalamus rajoittuu osittain erikoistuneisiin aivoalueisiin, joilta puuttuu tehokas veri-aivoeste; näissä kohdissa kapillaarien endoteeli on fenestroitunut, jotta suuretkin proteiinit ja muut molekyylit pääsevät vapaasti läpi. Jotkin näistä alueista ovat neurosekretion paikkoja – neurohypofyysi ja median eminence. Toiset ovat kuitenkin kohtia, joissa aivot ottavat näytteitä veren koostumuksesta. Kaksi näistä paikoista, SFO (subfornical organ) ja OVLT (organum vasculosum of the lamina terminalis) ovat niin sanottuja circumventrikulaarisia elimiä, joissa neuronit ovat läheisessä kosketuksessa sekä veren että CSF:n kanssa. Nämä rakenteet ovat tiheästi vaskularisoituneita, ja niissä on osmoreettisia ja natriumia vastaanottavaisia neuroneja, jotka säätelevät juomista, vasopressiinin vapautumista, natriumin erittymistä ja natriumin ruokahalua. Ne sisältävät myös neuroneja, joilla on reseptorit angiotensiinille, eteisnatriureettiselle tekijälle, endoteliinille ja relaksiinille, joista jokainen on tärkeä neste- ja elektrolyyttitasapainon säätelyssä. OVLT:n ja SFO:n neuronit heijastuvat supraoptiseen ytimeen ja paraventrikulaariseen ytimeen sekä myös preoptisiin hypotalamuksen alueisiin. Circumventrikulaariset elimet voivat myös olla interleukiinien vaikutuskohde, joka saa aikaan sekä kuumeen että ACTH:n erityksen, vaikutusten kautta paraventrikulaarisiin neuroneihin.
Ei ole selvää, miten kaikki hypotalamuksen toimintaan vaikuttavat peptidit saavat tarvittavan pääsyn. Prolaktiinin ja leptiinin tapauksessa on näyttöä aktiivisesta imeytymisestä suonikalvopleksuksessa verestä aivo-selkäydinnesteeseen (CSF). Joillakin aivolisäkkeen hormoneilla on negatiivinen takaisinkytkentä hypotalamuksen eritykseen; esimerkiksi kasvuhormoni vaikuttaa takaisin hypotalamukseen, mutta ei ole selvää, miten se pääsee aivoihin. Myös prolaktiinin sentraalisista vaikutuksista on näyttöä.
Tulosten mukaan kilpirauhashormoni (T4) päätyy hypotalamuksen gliasoluihin infundibulaarisessa ytimessä/mediaani-eminenssissä ja muuttuu siellä T3:ksi tyypin 2 deiodinaasin (D2) avulla. Tämän jälkeen T3 kulkeutuu tyrotropiinia vapauttavan hormonin (TRH) tuottaviin neuroneihin paraventrikulaarisessa ytimessä. Näistä neuroneista on löydetty kilpirauhashormonireseptoreita, mikä osoittaa, että ne ovat todellakin herkkiä T3-ärsykkeille. Lisäksi nämä neuronit ilmentävät MCT8:a, kilpirauhashormonin kuljettajaa, mikä tukee teoriaa siitä, että T3 kulkeutuu niihin. T3 voisi sitten sitoutua näissä neuroneissa olevaan kilpirauhashormonireseptoriin ja vaikuttaa kilpirauhashormonia vapauttavan tyrotropiinia vapauttavan hormonin tuotantoon, jolloin kilpirauhashormonituotantoa säädellään.
Hypotalamus toimii eräänlaisena elimistön termostaattina. Se asettaa halutun ruumiinlämpötilan ja stimuloi joko lämmöntuotantoa ja -pidätystä veren lämpötilan nostamiseksi korkeampaan asetukseen tai hikoilua ja verisuonten laajentumista veren jäähdyttämiseksi matalampaan lämpötilaan. Kaikki kuumeet johtuvat hypotalamuksen kohonneesta asetuksesta; muusta syystä johtuva kohonnut ruumiinlämpö luokitellaan hypertermiaksi. Harvoin hypotalamuksen suora vaurioituminen, esimerkiksi aivohalvauksen seurauksena, aiheuttaa kuumetta; tätä kutsutaan joskus hypotalamuskuumeeksi. Tavallisempaa on kuitenkin, että tällainen vaurio aiheuttaa epänormaalin alhaisen ruumiinlämmön.
SteroiditEdit
Hypotalamus sisältää neuroneja, jotka reagoivat voimakkaasti steroideihin ja glukokortikoideihin – (lisämunuaisen steroidihormoneihin, joita vapautuu vastauksena ACTH:lle). Se sisältää myös erikoistuneita glukoosille herkkiä neuroneja (kaarevassa ytimessä ja ventromediaalisessa hypotalamuksessa), jotka ovat tärkeitä ruokahalun kannalta. Preoptisella alueella on lämpöherkkiä neuroneja; nämä ovat tärkeitä TRH:n eritykselle.
NeuralEdit
Oksitosiinin eritystä vasteena imemis- tai vagino-cervikaaliseen stimulaatioon välittävät jotkin näistä reiteistä; vasopressiinin eritystä vasteena kardiovaskulaarisiin ärsykkeisiin, jotka syntyvät kaulavaltimoiden rungon ja aortan kaaren kemoreseptoreista sekä matalapaineisten eteisvolyymin reseptoreista, välittävät toiset. Rotilla emättimen stimulaatio aiheuttaa myös prolaktiinin eritystä, ja tämä johtaa hedelmättömän parittelun jälkeiseen näennäisraskauteen. Kaniinilla yhdyntä saa aikaan refleksinomaisen ovulaation. Lampaalla kohdunkaulan stimulaatio korkeiden estrogeenipitoisuuksien läsnä ollessa voi saada aikaan neitsytuoreen uuhen äidin käyttäytymisen. Kaikki nämä vaikutukset välittyvät hypotalamuksesta, ja tiedot kulkevat pääasiassa aivorunkoon välittyvien selkäydinreittien kautta. Nännien stimulaatio stimuloi oksitosiinin ja prolaktiinin vapautumista ja vaimentaa LH:n ja FSH:n vapautumista.
Kardiovaskulaariset ärsykkeet välittyvät vagushermon kautta. Vagus välittää myös monenlaista viskeraalista informaatiota, mukaan lukien esimerkiksi mahalaukun paisumisesta tai tyhjenemisestä johtuvat signaalit, jotka tukahduttavat tai edistävät ruokailua viestittämällä leptiini- tai gastriinin vapautumisesta. Nämäkin tiedot saapuvat hypotalamukseen aivorungossa sijaitsevien releiden kautta.
Lisäksi hypotalamuksen toiminta reagoi ja sitä säätelevät kaikkien kolmen klassisen monoamiinivälittäjäaineen, noradrenaliinin, dopamiinin ja serotoniinin (5-hydroksitryptamiinin) pitoisuudet niissä hermoradoissa, joista se saa innervaatiota. Esimerkiksi locus coeruleuksesta peräisin olevilla noradrenergisillä syötteillä on merkittäviä sääteleviä vaikutuksia kortikotropiinia vapauttavan hormonin (CRH) tasoihin.
Ruokailun säätelyTiedosto
| Ruokailua lisäävät peptidit . käyttäytymistä |
Peptidit, jotka vähentävät ruokintakäyttäytymistä |
|---|---|
| Ghrelin | Leptin |
| Neuropeptidi Y | (α,β,γ)-Melanosyyttejä stimuloivat hormonit |
| Agoutiin liittyvä peptidi | Kokaiinin ja amfetamiinin säätelemät transkriptiopeptidit |
| Orexiinit (A,B) | Kortikotropiinia vapauttava hormoni |
| Melaniinikonsentroiva hormoni | Kolekystokiniini |
| Galaniini | Insuliini |
| > | glukagoni-like peptide 1 |
Hypotalamuksen ventromediaalisen ytimen äärimmäinen lateraalinen osa vastaa ruoan saannin ohjauksesta. Tämän alueen stimulaatio aiheuttaa lisääntynyttä ruoan saantia. Tämän alueen molemminpuolinen vaurio aiheuttaa ruoan saannin täydellisen loppumisen. Ytimen mediaalisilla osilla on ohjaava vaikutus lateraaliseen osaan. Ventromediaalisen ytimen mediaalisen osan bilateraalinen vaurio aiheuttaa hyperfagiaa ja eläimen lihomista. Ventromediaalisen ytimen lateraalisen osan myöhempi leesio samassa eläimessä saa aikaan ruoanoton täydellisen loppumisen.
Tähän säätelyyn liittyy erilaisia hypoteeseja:
- Lipostaattinen hypoteesi: Tämän hypoteesin mukaan rasvakudos tuottaa humoraalisen signaalin, joka on suhteessa rasvan määrään ja vaikuttaa hypotalamukseen vähentääkseen ruoan saantia ja lisätäkseen energiantuottoa. On ollut ilmeistä, että hormoni leptiini vaikuttaa hypotalamukseen vähentääkseen ruoan saantia ja lisätäkseen energiantuottoa.
- Gutpeptidihypoteesi: Ruoansulatuskanavan hormonien, kuten Grp:n, glukagonien, CCK:n ja muiden väitetään estävän ruoan saantia. Ruoan joutuminen ruoansulatuskanavaan laukaisee näiden hormonien vapautumisen, jotka vaikuttavat aivoihin tuottaakseen kylläisyyden. Aivoissa on sekä CCK-A- että CCK-B-reseptoreita.
- Glukostaattihypoteesi: Ventromediaalisissa ytimissä sijaitsevan kylläisyyskeskuksen toimintaa ohjaa todennäköisesti glukoosin käyttö neuroneissa. On esitetty, että kun niiden glukoosin hyväksikäyttö on vähäistä ja näin ollen kun niiden kautta kulkeva arteriovenoosinen verenglukoosiero on pieni, neuronien kautta kulkeva aktiivisuus vähenee. Näissä olosuhteissa ravintokeskuksen aktiivisuus on hallitsematonta ja yksilö tuntee nälkää. Ruoan saanti lisääntyy nopeasti laskimonsisäisellä 2-deoksiglukoosin antamisella, jolloin glukoosin käyttö soluissa vähenee.
- Termostaattinen hypoteesi: Tämän hypoteesin mukaan ruumiinlämmön lasku alle tietyn asetusarvon stimuloi ruokahalua, kun taas nousu asetusarvon yläpuolelle estää ruokahalua.
Pelon prosessointiMuutos
Hypotalamuksen mediaalinen vyöhyke on osa virtapiiriä, joka kontrolloi motivoitunutta käyttäytymistä, kuten puolustuskäyttäytymistä. Fos-merkinnän analyysit osoittivat, että sarja ytimiä ”käyttäytymisen ohjauspylväässä” on tärkeä synnynnäisen ja ehdollistetun puolustuskäyttäytymisen ilmentymisen säätelyssä.
Antipredatorinen puolustuskäyttäytyminen
Altistuminen petoeläimelle (kuten kissalle) herättää laboratoriojyrsijöissä puolustuskäyttäytymistä silloinkin, kun eläin ei ole koskaan ollut alttiina kissalle. Hypotalamuksessa tämä altistuminen aiheuttaa Fos-merkittyjen solujen lisääntymistä hypotalamuksen etuosan ytimessä, ventromediaalisen ytimen dorsomediaalisessa osassa ja premammillaarisen ytimen ventrolateraalisessa osassa (PMDvl). Premammillaarisella ytimellä on tärkeä rooli petoeläimiin kohdistuvan puolustuskäyttäytymisen ilmentymisessä, sillä tämän ytimen vauriot poistavat puolustuskäyttäytymisen, kuten jäätymisen ja pakenemisen. PMD ei moduloi puolustuskäyttäytymistä muissa tilanteissa, sillä tämän ytimen vaurioilla oli minimaalinen vaikutus shokin jälkeisiin jäätymisarvoihin. PMD:llä on tärkeät yhteydet dorsaaliseen periaqueductal greyyn, joka on tärkeä rakenne pelon ilmaisussa. Lisäksi eläimet osoittavat riskinarviointikäyttäytymistä ympäristöön, joka on aiemmin liitetty kissaan. Fos-merkittyjen solujen analyysi osoitti, että PMDvl on hypotalamuksen aktivoitunein rakenne, ja inaktivointi muscimolilla ennen kontekstille altistumista poistaa puolustuskäyttäytymisen. Näin ollen hypotalamuksella, pääasiassa PMDvl:llä, on tärkeä rooli synnynnäisen ja ehdollistetun puolustuskäyttäytymisen ilmentymisessä saalistajaa kohtaan.
Sosiaalinen tappio
Kuten myös hypotalamuksella on rooli sosiaalisessa tappiossa: Mediaalisen vyöhykkeen ytimet mobilisoituvat myös aggressiivisen lajitoverin kohtaamisen aikana. Lyödyn eläimen Fos-tasot kohoavat sukupuolidimorfisissa rakenteissa, kuten mediaalisessa preoptisessa ytimessä, ventromediaalisen ytimen ventrolateraalisessa osassa ja ventraalisessa premammilaarisessa ytimessä. Tällaiset rakenteet ovat tärkeitä muussa sosiaalisessa käyttäytymisessä, kuten seksuaalisessa ja aggressiivisessa käyttäytymisessä. Lisäksi myös premammillaarinen ydin mobilisoituu, dorsomediaalinen osa mutta ei ventrolateraalinen osa. Tämän ytimen vauriot poistavat passiivisen puolustuskäyttäytymisen, kuten jäätymisen ja ”selällään” -asennon.