ホルモン分泌編
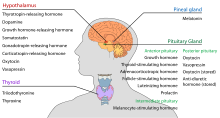
視床下部は中枢神経内分泌機能を持ち、最も顕著なのは、その制御により、下垂体の前部が様々な内分泌腺や器官の制御をしていることである。 放出ホルモン(放出因子とも呼ばれる)は視床下部の核で産生され、軸索に沿って正中隆起または下垂体後葉に運ばれ、そこで貯蔵され、必要に応じて放出される。
下垂体前葉
視床下部-腺下垂体軸では、放出ホルモンは、下垂体向性ホルモンまたは下垂体ホルモンとしても知られ、視床下部の延長である中央隆起から下垂体門系に放出され、下垂体前葉に運ばれて腺下垂体ホルモンの分泌に対する制御機能を発揮する。 これらの下垂体刺激ホルモンは、視床下部の脳室周囲に存在する傍細胞性神経分泌細胞によって刺激される。 下垂体ホルモンは第3脳室の毛細血管に放出された後、視床下部-下垂体門脈循環と呼ばれる経路で移動する。 下垂体前葉に到達すると、これらのホルモンは下垂体細胞の表面に存在する特定の受容体に結合します。 この結合によってどの細胞が活性化されるかによって、下垂体は血流の残りの部分にホルモンの分泌を開始したり停止したりする。
| 分泌ホルモン | 略称 | 産生 | 効果 | |
|---|---|---|---|---|
| TRH、TRF, or PRH | 室傍核の神経分泌細胞 | 下垂体前葉からの甲状腺刺激ホルモン(TSH)放出を刺激(主に) 下垂体前葉からのプロラクチン放出を刺激 |
||
| Corticotropin-> | 下垂体前葉の細胞からの神経分泌細胞放出ホルモン | CRH or CRF | 室傍核の神経分泌細胞 | 下垂体前葉からの副腎皮質刺激ホルモン(ACTH)放出を刺激 |
| Dopamine (プロラクチン抑制ホルモン) |
DA or PIH | Dopamine neurons of arcuate nucleus | Inhibit proactin release from anterior pituitary | |
| Growth->Dopamine neuron from anterior pituitary | GHRH | 弧状核の神経内分泌ニューロン | Stimulate growth-?下垂体前葉からのホルモン(GH)放出 | |
| Gonadotropin-releasing hormone | GnRH or LHRH | Preoptic areaの神経内分泌細胞 | Stimulate follicle-> | Stimulate follicle-> |
| ソマトスタチン (成長ホルモン抑制ホルモン) |
SS, GHIH, or SRIF | 室周囲核の神経内分泌細胞 | 下垂体前葉からの成長ホルモン(GH)放出を抑制する 下垂体前葉からの甲状腺刺激ホルモン(TSH)放出を(適度に)抑制する |
中央隆起からの他のホルモンとしてバゾプレシンが分泌される。 オキシトシン、ニューロテンシン。
下垂体後葉
視床下部-神経下垂体軸では、実際には視床下部の延長である下垂体後葉から神経下垂体ホルモンが循環系に放出される。
| 分泌ホルモン | 略称 | 産生 | 効果 |
|---|---|---|---|
| Oxytocin | OXY or OXT | 室傍核と上丘の拡大細胞神経分泌性セルがある 核 | 子宮収縮 授乳(letdown reflex) |
| バソプレシン (抗利尿ホルモン) |
ADH or AVP | 室傍核の細胞および傍細胞神経内分泌系細胞。 視索上核の大細胞 | 腎臓の遠位尿細管と集合管の細胞の水に対する透過性が増加し、水の再吸収と濃縮尿の排泄が可能になる |
また、特定の皮膚疾患や皮膚の恒常性と、視床下部-下垂体-副腎軸(HPA)ホルモンが関連していることが知られています。
刺激編集
視床下部は、多くのホルモンおよび行動の概日リズム、神経内分泌出力の複雑なパターン、複雑な恒常性維持機構、および重要な行動を調整している。 したがって、視床下部は多くの異なるシグナルに反応しなければならないが、その中には外部で発生するものと内部で発生するものがある。 視床下部または大脳皮質で発生するデルタ波信号が放出ホルモンの分泌に影響を与え、GHRH とプロラクチンが刺激される一方、TRH は抑制されます。
視床下部は次のことに反応します:
- 光。 フェロモンなどの嗅覚刺激
- 生殖腺ステロイドや副腎皮質ホルモンなどのステロイド
- 特に心臓、腸神経系(消化管の)、生殖管から生じる神経伝達情報
- 視床下部の反応:光:日長と光周期(概日リズムおよび季節リズムを調節するため)
- 視覚刺激(生理的な刺激、生理的な刺激)。
- 自律神経入力
- レプチン、グレリン、アンジオテンシン、インスリン、下垂体ホルモン、サイトカイン、グルコースの血漿濃度および浸透圧など、血液から生じる刺激。
- ストレス
- 体温を上昇させて微生物を侵入させ、体のサーモスタットを上にリセットする。
嗅覚刺激編集
多くの種で有性生殖や神経内分泌機能に嗅覚刺激が重要であるとされる。 例えば、妊娠中のマウスが交尾後の重要な時期に「見知らぬ」オスの尿にさらされると、妊娠は失敗に終わります(ブルース効果)。 このように、雌のマウスは性交中に相手に対する正確な「嗅覚記憶」を形成し、それが数日間持続するのである。 フェロモンによる合図は、多くの種で発情の同期化を助ける。女性では、月経の同期化もフェロモンによる合図から生じるかもしれないが、ヒトにおけるフェロモンの役割については議論があるところである。 視床下部は、有効な血液脳関門を持たない特殊な脳領域に一部囲まれています。これらの部位の毛細血管内皮は、大きなタンパク質や他の分子でさえ自由に通過できるように柵状になっています。 これらの部位には、神経下垂体や正中隆起といった神経分泌の場もある。 しかし、それ以外の部位は、脳が血液の組成をサンプリングする部位である。 これらの部位のうち、SFO(subfornical organ)とOVLT(organum vasculosum of the lamina terminalis)はいわゆるcircumventricular organで、神経細胞が血液とCSFの両方に密接に接触している場所である。 これらの構造体は高密度に血管が張り巡らされており、飲水、バソプレシン放出、ナトリウム排泄、ナトリウム食欲を制御するオスモレセプターおよびナトリウムレセプターニューロンを含んでいる。 また、アンジオテンシン、心房性ナトリウム利尿因子、エンドセリンおよびリラキシンに対する受容体を持つニューロンも含まれており、これらはそれぞれ体液および電解質バランスの調節において重要である。 OVLT と SFO のニューロンは、視床上部の核と室傍核、および視床下部前部へ投射しています。 また、室傍ニューロンへの影響を介して、発熱とACTH分泌の両方を誘発するインターロイキンの作用部位である可能性もある。
視床下部の活動に影響を与えるすべてのペプチドが、どのようにして必要なアクセスを獲得しているかは明らかでない。 プロラクチンやレプチンの場合、血液から脳脊髄液(CSF)への脈絡叢での活発な取り込みが証明されている。 下垂体ホルモンの中には、視床下部の分泌に負のフィードバック影響を及ぼすものがある;例えば、成長ホルモンは視床下部にフィードバックするが、それがどのように脳に入るかは不明である。 甲状腺ホルモン(T4)は視床下部の下垂体核/正中隆起のグリア細胞に取り込まれ、ここで2型デイオジナーゼ(D2)によりT3に変換されるとする研究結果もある。 その後、T3は室傍核のチロトロピン放出ホルモン(TRH)産生ニューロンへ輸送される。 この神経細胞には甲状腺ホルモン受容体が見つかっており、実際にT3刺激に敏感であることが示されている。 さらに、これらのニューロンには甲状腺ホルモン輸送体であるMCT8が発現しており、T3が輸送されるという説が支持された。 8306>
視床下部は体のサーモスタットのようなものである。 望ましい体温を設定し、血温を高く設定するために熱産生と保温を、あるいは血温を低くするために発汗と血管拡張を刺激するのである。 すべての発熱は、視床下部の設定温度の上昇に起因する。その他の原因による体温の上昇は、高体温症に分類される。 まれに、脳卒中などで視床下部が直接損傷すると発熱することがあり、これを視床下部熱と呼ぶこともあります。 視床下部には、ステロイドおよびグルココルチコイド(ACTHに反応して放出される副腎のステロイドホルモン)に強く反応する神経細胞がある。 また、食欲に重要なグルコース感受性の特殊なニューロン(視床下部弓状核と視床下部内側)も含まれています。
NeuralEdit
乳汁分泌や膣頸部刺激に応じたオキシトシン分泌は、これらの経路の一部によって媒介される。頸動脈体および大動脈弓の化学受容体、および低圧心房容量受容体に生じる心血管刺激に応じたバソプレシン分泌は、他の経路によって媒介される。 ラットでは、膣の刺激によってもプロラクチンが分泌され、不妊交配後の擬似妊娠につながる。 ウサギでは、交尾により反射的に排卵が誘発される。 ヒツジでは、高濃度のエストロゲンの存在下で子宮頸管を刺激すると、処女の雌羊に母性行動を誘発することができます。 これらの作用はすべて視床下部が仲介し、情報は主に脳幹を中継する脊髄経路によって運ばれる。 乳首を刺激すると、オキシトシンやプロラクチンの放出が促され、LHやFSHの放出が抑制される。
心血管系の刺激は迷走神経によって運ばれる。 迷走神経はまた、例えば胃の膨張や空虚から生じる信号など、さまざまな内臓情報を伝え、それぞれレプチンやガストリンの放出を合図に、摂食を抑制したり促進したりする。 8306>
さらに視床下部の機能は、視床下部が神経支配を受けている経路の3つの古典的なモノアミン神経伝達物質、ノルアドレナリン、ドーパミン、セロトニン(5-ヒドロキシトリプタミン)のレベルに反応し、それによって調節される。 例えば、ノルアドレナリンの入力は、コルチコトロピン放出ホルモン(CRH)のレベルを調節する重要な作用がある。
摂食の制御Edit
| 摂食を促進するペプチド 摂食 行動 |
摂食行動 |
|---|---|
| グレリン | レプチン | (α.蝙九髟キ蟆コ隱ソ謨) |
| アグーチ関連ペプチド | コカインおよびアンフェタミン制御転写ペプチド | コルチコトロピン放出ホルモン | メラニン濃度ホルモン | コレシストキニン | ガラニン | インシュリン | グルカゴン- | インシュラングリッパー |
視床下部の腹内側核の極外側は、食物摂取の制御に関与している。 この部位を刺激すると、食物摂取量が増加する。 この部位の両側の病変は食物の摂取を完全に停止させる。 核の内側は外側を制御する作用がある。 腹内側核の内側部分の両側病変は動物の食欲亢進と肥満を引き起こす。 さらに、同じ動物で腹内側核の外側部分を損傷すると、食物の摂取が完全に停止する。
この制御に関するさまざまな仮説がある。 この仮説は、脂肪組織が脂肪量に比例した体液性シグナルを生成し、視床下部に作用して食物摂取量を減少させ、エネルギー出力を増加させるとするものである。 レプチンというホルモンが視床下部に作用して、食物摂取量を減少させ、エネルギー出力を増加させることが明らかになっている。
Fear processing 編集
視床下部の内側領域は防衛行動などの意欲的行動を制御する回路の一部であり、視床下部は防衛行動を制御している。 Fos標識の解析から、「行動制御コラム」にある一連の核が、生得的な防御行動と条件付けられた防御行動の発現を制御する上で重要であることが示された。
反捕食性防御行動
実験用げっ歯類では、ネコなどの捕食者にさらされると、ネコと接触したことがない動物でも防御行動を誘発させることが分かっている。 視床下部では、この曝露により視床下部前核、腹内側核の背内側部分、乳腺前核の腹外側部分(PMDvl)でFos標識細胞が増加した。 乳房前核は捕食者に対する防御行動の発現に重要な役割を担っており、この核の病変は凍りつきや飛翔などの防御行動を消失させるからです。 この核の病変は、ショック後の凍りつきスコアにほとんど影響を及ぼさないことから、PMDは他の状況での防衛行動には影響を及ぼさないことがわかりました。 PMDは、恐怖の発現に重要な構造である背側枝状回廊灰白と重要な結合を有している。 さらに、動物は、以前猫と関連した環境に対するリスク評価行動を示す。 Fos標識細胞解析の結果、PMDvlは視床下部で最も活性化された構造であり、文脈に暴露する前にムシモールで不活性化すると防衛行動が消失することが示された。 8306>社会的敗北<4927>同様に、視床下部は社会的敗北に関与している。 視床下部も攻撃的な相手と遭遇すると、内側核が動員される。 敗北した動物は内側視蓋前核、腹内側核の腹外側部分、腹側乳腺前核など性差のある構造でFosレベルが上昇した。 これらの構造は、性的行動や攻撃的行動など、他の社会的行動においても重要である。 さらに、乳腺前核も動員され、背内側部分は動員されるが、腹外側部分は動員されない。 この核を損傷すると、凍りつきや「仰向け」姿勢などの受動的な防衛行動が消失する
。