Introduction
Gram-positieve bacteriën van de genera Bacillus en Clostridium kunnen zich differentiëren tot een endospore (spore), een metabolisch rustende cel die wordt geproduceerd als reactie op omstandigheden die celgroei niet toelaten (McKenney et al., 2012). Eenmaal in het milieu vrijgegeven, overleeft de spore extreem lang in zijn slapende toestand en is hij bestand tegen een groot aantal stressfactoren, zoals hoge temperaturen, uitdroging, afwezigheid van voedingsstoffen en de aanwezigheid van giftige chemicaliën (McKenney et al., 2012). De spore in ruststadium is echter in staat om de omgeving voortdurend te monitoren en te reageren op de aanwezigheid van water en voedingsstoffen door te ontkiemen en een vegetatieve cel te vormen die kan groeien en sporuleren (McKenney et al., 2012). Weerstand tegen niet-fysiologische omstandigheden is deels te danken aan de oppervlaktestructuren van de sporen. Bij Bacillus subtilis, het modelsysteem voor sporenvormers, is het sporenoppervlak georganiseerd in een meerlagige laag en in een korst, een recent ontdekte laag die de sporenlaag omringt (McKenney et al., 2012). B. subtilis-sporen zijn negatief geladen (Huang et al., 2010; Pesce et al., 2014) en hebben een relatieve hydrofobiciteit die deels het gevolg is van de glycosylering van sommige spore-oppervlak-eiwitten (Cangiano et al., 2014; Rusciano et al., 2014). Bij verschillende Bacillus- en Clostridium-soorten, waaronder B. cereus, B. anthracis, B. megaterium en C. difficile, is de buitenste sporenstructuur het exosporium, een morfologisch onderscheiden laag bestaande uit eiwitten en glycoproteïnen die de mantel omgeeft (Díaz-González et al., 2015; Manetsberger et al, 2015b; Stewart, 2015).
De bacteriële spore is voorgesteld als een platform om heterologe eiwitten weer te geven, met potentiële toepassingen variërend van de ontwikkeling van mucosale vaccins tot herbruikbare biokatalysatoren, diagnostische hulpmiddelen en bioremediëringsapparaten (Knecht et al., 2011; Isticato en Ricca, 2014; Ricca et al., 2014). De opmerkelijke en goed gedocumenteerde resistentie van de spore (McKenney et al., 2012), de vatbaarheid van verschillende sporevormende soorten voor genetische manipulatie (Harwood and Cutting, 1990) en de veiligheid van verschillende soorten (Cutting, 2011; Baccigalupi et al., 2015) ondersteunen het gebruik van de spore als een weergave- en afgiftesysteem. Er zijn twee strategieën ontwikkeld om heterologe eiwitten op het spore-oppervlak weer te geven. Een recombinante strategie, gebaseerd op de constructie van genfusies tussen DNA dat codeert voor een geselecteerd spore-oppervlak-eiwit en DNA dat codeert voor het weer te geven eiwit, is in de loop der jaren gebruikt om een verscheidenheid aan heterologe eiwitten weer te geven (Isticato en Ricca, 2014). Een niet-recombinante benadering, gebaseerd op de spontane adsorptie tussen gezuiverde sporen en gezuiverde eiwitten, is ook gebruikt om verschillende enzymen en antigenen weer te geven (Ricca et al., 2014). De moleculaire details die de adsorptie van sporen regelen, zijn nog niet volledig opgehelderd. Het is bekend dat de adsorptie efficiënter is wanneer de pH van de bindingsbuffer zuur is (pH 4) (Huang et al., 2010; Sirec et al., 2012) en dat waarschijnlijk een combinatie van elektrostatische en hydrofobe interacties bij de interactie betrokken is (Huang et al., 2010; Sirec et al., 2012). Het is ook bekend dat mutantsporen met sterk veranderde sporenoppervlakken efficiënter dan isogene wild-type sporen interageren met modeleiwitten (Sirec et al., 2012; Donadio et al, 2016).
Hier hebben we een fluorescerend eiwit gebruikt, de monomere vorm van het Rood Fluorescerend Eiwit (mRFP) van het koraal Discosoma sp. (Campbell et al., 2002), om te evalueren of sporen van B. megaterium in staat zijn tot interactie met en adsorptie van een model heteroloog eiwit. B. megaterium omvat een aantal morfologisch verschillende stammen die de ongewoon grote afmetingen van zowel cellen (lengte tot 4 μm en een diameter van 1,5 μm) als sporen (lengte tot 3 μm en een diameter van 1 μm) delen (Di Luccia et al., 2016). Sporen van sommige stammen van B. megaterium zijn omgeven door een exosporium, en aangezien tot nu toe alleen sporen van soorten die geen exosporium hebben als adsorptieplatforms zijn beschouwd, zijn er geen gegevens beschikbaar over de impact van het exosporium in de interactie met heterologe eiwitten.
De QM B1551-stam is de best gekarakteriseerde stam van B. megaterium. Deze stam draagt ongeveer 11% van zijn genoom op zeven inheemse plasmiden (Rosso en Vary, 2005; Vary et al., 2007; Eppinger et al., 2011), waarvan er twee – pBM500 en pBM600 – zijn geïdentificeerd als dragers van genen die essentieel zijn voor de vorming van het kenmerkende “walnootvormige” exosporium van deze stam (Manetsberger et al., 2015a). De eiwitsamenstelling van het QM B1551 exosporium is tot nu toe slecht gekarakteriseerd, met slechts een paar genen die coderen voor orthologs van erkende exosporium eiwitten in sporen van andere soorten die door genomische analyses zijn geïdentificeerd. Hiertoe behoren genen die coderen voor BclA nap-eiwitten, die een gelokaliseerde nap vormen in sporen van B. megaterium QM B1551, plus een ortholoog van het BxpB-eiwit dat de basale laag van het exosporium vormt in sporen van de familie B. cereus. Het laatste lijkt een andere rol te vervullen in B. megaterium QM B1551 sporen, aangezien een null mutant stam een ogenschijnlijk normaal exosporium behield (Manetsberger et al, 2015a).
In dit artikel presenteren we gegevens die aantonen dat sporen van B. megaterium QM B1551 mRFP efficiënt kunnen adsorberen, en bewijs leveren dat eiwitmoleculen in staat zijn om de doorlaatbaarheidsbarrière die door het exosporium wordt gepresenteerd te doorkruisen om zich in de intercoatingruimte te lokaliseren.
Materialen en methoden
Bacteriestammen, sporen, en RFP productie
De B. megaterium stammen gebruikt in deze studie zijn QM B1551 en zijn plasmide-loze derivaat PV361 (Rosso en Vary, 2005). De in deze studie gebruikte B. subtilis stam was PY79 (Youngman et al., 1984). Sporulatie van alle Bacillus-stammen werd geïnduceerd met de uitputtingsmethode (Cutting en Vander Horn, 1990). Na 30 uur groei in Difco Sporulatie (DS) medium bij 37°C onder krachtig schudden werden sporen verzameld en gezuiverd zoals beschreven door Nicholson en Setlow (1990) met behulp van nachtelijke incubatie in H2O bij 4°C om resterende sporangiale cellen te lyseren. Het aantal verkregen gezuiverde sporen werd gemeten door directe telling met een Bürker-kamer onder een optische microscoop (Olympus BH-2 met 40× lens).
Voor mRFP-productie werden cellen van Escherichia coli-stam RH161 (Donadio et al., 2016), die de expressievector pRSET-A dragen met een in-frame fusie van het 5′ einde van de rfp coderende regio tot zes histidine codons onder de transcriptionele controle van een T7 promotor, werden gekweekt gedurende 18 uur bij 37 ° C in 100 ml auto-inductie medium om het heterologe eiwit tot expressie te brengen (Isticato et al., 2010). Het His6-getagde RFP-eiwit werd gezuiverd onder natieve omstandigheden met behulp van een His-Trap kolom zoals aanbevolen door de fabrikant (GE Healthcare Life Science). Gezuiverd eiwit werd ontzout met behulp van een PD10 kolom (GE Healthcare Life Science) om hoge NaCl en imidazool concentraties te verwijderen.
Adsorptiereactie
Niet anders gespecificeerd 5 ug gezuiverd recombinant mRFP werd toegevoegd aan een suspensie van sporen (5 × 108) in 50 mM natriumcitraat pH 4,0 bij 25 ° C in een eindvolume van 200 pi. Na 1 uur incubatie werd het bindingsmengsel gecentrifugeerd (10 min bij 13.000 g) om mRFP gebonden sporen in de pellet te fractioneren van vrije mRFP in het supernatant.
Western en Dot-Blot Analysis
Sporenpellets van adsorptiereacties werden geresuspendeerd in 40 pi sporevachtextractiebuffer (Nicholson en Setlow, 1990; Giglio et al, 2011), gedurende 1 uur geïncubeerd bij 68°C om sporencoatingseiwitten op te lossen en op een 12% SDS-PAGE-gel geladen. De eiwitten werden vervolgens geëlektrolyseerd naar nitrocellulose filters (Amersham Pharmacia Biotech) en gebruikt voor Western blot analyse zoals eerder gerapporteerd (Isticato et al., 2008) met behulp van monoklonale mRFP-herkennend anti-His antilichaam (Sigma). Een kwantitatieve bepaling van de hoeveelheid mRFP werd verkregen door dot blot experimenten vergelijking van seriële verdunningen van gezuiverd mRFP en binding assay supernatant. De filters werden vervolgens gevisualiseerd met de ECL-prime methode (Amersham Pharmacia Biotech) en onderworpen aan densitometrische analyse met Quantity One 1-D Analysis Software (Bio-Rad). Dot blot en relatieve densitometrische analyses werden drie keer uitgevoerd om de significantie van de resultaten te verifiëren.
Fluorescentie en immunofluorescentie microscopie
Post-adsorptie sporen werden geresuspendeerd in 50 ul van 1x PBS pH 4,0 en 5 ul van de suspensie geplaatst op microscoopglaasjes en bedekt met een dekglaasje vooraf behandeld gedurende 30 s met poly-L-lysine (Sigma). Immunofluorescentie werd uitgevoerd zoals beschreven door Isticato et al. (2013), met de volgende wijzigingen: 2,0 × 106 RFP-adsorbed sporen van QM B1551 en PV361 B. megaterium stammen werden voorbehandeld met 1% runderserumalbumine (BSA) – 1x PBS pH 4,0 gedurende 30 min voorafgaand aan 2 h-incubatie bij 4 ° C met de anti-polyHistidine antilichamen (muis; Sigma) verdund 1:20 in 1x PBS pH 4,0-1% BSA. Ter controle van de specificiteit van deze techniek werden niet-geadsorbeerde sporen direct behandeld met anti-His antilichamen. Na driemaal wassen werden de monsters geïncubeerd met een 64-voudig verdund anti-muis secundair antilichaam geconjugeerd met fluoresceïne-isothiocyanaat, FITC (Santa Cruz Biotechnology, Inc.) en viermaal gewassen met 1x PBS pH 4.0. Gewassen monsters werden geresuspendeerd in 20 ul van 1x PBS pH 4,0 en 10 ul werden geanalyseerd. Alle monsters werden waargenomen met een Olympus BX51 fluorescentiemicroscoop uitgerust met een 100 × UPlan F1 olie objectief; U-MNG of U-MWIBBP kubus-filters werden gebruikt om de rode fluorescentie-emissie van mRFP of de groene emissie van FITC-geconjugeerde antilichamen te detecteren, respectievelijk. De belichtingstijden lagen tussen 200 en 3000 ms. Beelden werden vastgelegd met een Olympus DP70 digitale camera en verwerkt met Image Analysis Software (Olympus) voor kleine aanpassingen van helderheid, contrast en kleurbalans (McCloy et al., 2014). ImageJ (v1.48, NIH) werd gebruikt om voor elke stam een omtrek rond 80 sporen te tekenen en de minimale, maximale en gemiddelde fluorescentiewaarden per pixel werden voor elke spore geregistreerd. Waarden van fluorescentie-intensiteit werden vervolgens weergegeven als box-plots met 5-95% betrouwbaarheidsintervallen (McCloy et al., 2014).
Statistische analyse
Resultaten van dot blot en fluorescentiemicroscopie analyse zijn de gemiddelden van drie onafhankelijke experimenten. Statistische significantie werd bepaald door de Student t-test, en het significantieniveau werd vastgesteld op P < 0.05.
Resultaten
mRFP van Discosoma sp. wordt geadsorbeerd door B. megaterium Spores
Om na te gaan of sporen van B. megaterium QM B1551 in staat waren mRFP te adsorberen, werd 5 μg van het gezuiverde eiwit (Materials and Methods) geïncubeerd met 5,0 × 108 gezuiverde sporen. De adsorptiereactie werd uitgevoerd in 50 mM natriumcitraat bij pH 4,0, zoals eerder beschreven (Sirec et al., 2012). Na de reactie werden sporen uitgebreid gewassen met 1x PBS pH 4,0, sporenoppervlak eiwitten werden geëxtraheerd zoals beschreven in Materials and Methods en geanalyseerd door western blotting met anti-polyHistidine-Peroxidase antilichaam (Sigma), dat de histidine-getagd N terminus van mRFP herkent. Zoals blijkt uit figuur 1A, werd mRFP geëxtraheerd uit sporen, wat aangeeft dat het werd geabsorbeerd tijdens de reactie en vervolgens vrijgegeven door de extractie behandeling. Om de efficiëntie van adsorptie te evalueren, analyseerden we de hoeveelheid mRFP links ongebonden, dat wil zeggen, post-geadsorbeerde sporen werden verzameld door centrifugatie en het supernatant serieel verdund en geanalyseerd door dot blotting (Figuur 1B). Een densitometrische analyse van de dot blot (Supplementary Table 1) toonde aan dat wanneer 5 ug mRFP werd gebruikt in de adsorptiereactie minder dan 1% werd links ongebonden, wat aangeeft dat ongeveer 99% van de heterologe eiwit werd geadsorbeerd aan B. megaterium spores.

FIGUUR 1. Bacillus megaterium QM B1551-sporen adsorberen mRFP. 5 × 108 sporen werden geïncubeerd met 5 μg gezuiverd mRFP en vervolgens werden de monsters gecentrifugeerd. (A) Spore oppervlakte-eiwitten werden geëxtraheerd uit de pelletfractie door SDS-DTT behandeling, gefractioneerd op SDS-PAGE en geanalyseerd door Western blot. Gezuiverde mRFP (20 μg) werd gebruikt als een marker. (B) Het supernatant, dat het ongebonden mRFP bevat, werd serieel verdund en geanalyseerd met dot blot (QM B1551). Seriële verdunningen van gezuiverd mRFP (mRFP) werden als standaard gebruikt. (C) Sporen geadsorbeerd met mRFP werden tweemaal gewassen (W1 en W2) met PBS buffer op pH 3,0 of pH 7,0, of met een 1M NaCl, 0,1% Triton X-100 oplossing. Seriële verdunningen van gezuiverd mRFP en ongebonden mRFP (UN-mRFP) werden gebruikt als standaard. Immuno-reacties in alle panelen werden uitgevoerd met mRFP-herkennend anti-His antilichaam geconjugeerd met mierikswortelperoxidase.
Om te analyseren of geadsorbeerde mRFP-moleculen stevig aan het sporenoppervlak waren gebonden, werden sporen na de adsorptiereactie twee keer gewassen met PBS-buffer bij pH 3,0 of pH 7,0, of met een 1M NaCl, 0,1% Triton X-100 oplossing, zoals eerder beschreven (Donadio et al., 2016). Zoals getoond in figuur 1C, en ondersteund door densitometrische analyse van de dot blot (Supplementary Table 2), de wasbeurten bij pH 3,0 veroorzaakte geen afgifte van de geadsorbeerde mRFP, terwijl de wasbeurten bij pH 7,0 of met 1M NaCl, 0,1% Triton X-100 veroorzaakte een minimale, minder dan 1%, afgifte van mRFP moleculen. Daarom suggereren de resultaten in figuur 1 dat mRFP efficiënt werd geadsorbeerd en stevig aan B. megaterium-sporen werd gebonden. Om te beoordelen of spore-geadsorbeerde mRFP moleculen hun fluorescentie eigenschappen behouden, voerden we een fluorescentie microscopie analyse. Zoals getoond in figuur 2, post-adsorptie reactie sporen werden geassocieerd met een sterke fluorescentie signaal zichtbaar rond het gehele spore oppervlak.
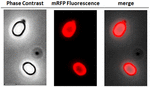
FIGUUR 2. Fluorescentiemicroscopie-analyse van B. megaterium QM B1551-mRFP-sporen. QM B1551 sporen geïncubeerd met mRFP (5 ug), en vervolgens gewassen, werden geanalyseerd met fluorescentiemicroscopie. Dezelfde microscopie veld werd waargenomen door fasecontrast en fluorescentie microscopie. Schaal bar 1 um. Het samenvoegingspaneel wordt gerapporteerd. De belichtingstijd was 200 ms.
Het exosporium heeft een essentiële rol in mRFP-adsorptie
Stam QM B1551 van B. megaterium bevat zeven inheemse plasmiden (Rosso en Vary, 2005; Eppinger et al., 2011) en plasmide-gecodeerde genen zijn essentieel voor exosporiumvorming (Manetsberger et al., 2015a). PV361 is een QM B1551-geharde stam die alle zeven plasmiden ontbeert en bijgevolg totaal geen exosporium heeft (Manetsberger et al., 2015a). We gebruikten sporen van stam PV361 om de rol van het exosporium in mRFP-adsorptie te analyseren. Parallel daaraan gebruikten we ook sporen van B. subtilis PY79 waarvan in een eerdere studie is aangetoond dat ze mRFP adsorberen (Donadio et al., 2016). Om de adsorptie-efficiëntie van sporen van de B. subtilis PY79 en B. megaterium QM B1551 en PV361 stammen te vergelijken, adsorbeerden we 5 μg gezuiverd mRFP met 5,0 × 108 sporen van elk van de drie stammen. Na de adsorptiereacties werden sporen verzameld door centrifugatie, eiwitten geëxtraheerd door SDS-DTT behandeling en geanalyseerd door western blotting met mRFP-herkennende anti-His antilichaam. Zoals blijkt uit figuur 3A, werd mRFP blijkbaar in grotere hoeveelheden geëxtraheerd uit sporen van QM B1551 dan uit sporen van de andere twee stammen. In een poging om deze schijnbare verschillen te kwantificeren, werd ongebonden mRFP uit de adsorptiereacties serieel verdund en geanalyseerd door dot blotting (figuur 3B). Een densitometrische analyse van de dot blot van figuur 3B (Supplementary Table 3) gaf aan dat PY79 en PV361 sporen geadsorbeerd ongeveer 90% van de totale mRFP, terwijl QM B1551 sporen geadsorbeerd bijna alle (meer dan 99%) gezuiverd mRFP.
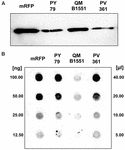
FIGUUR 3. Monomere rood fluorescerende proteïne-adsorptie aan sporen van B. megaterium QM B1551 en PV361 en B. subtilis PY79. 5 × 108 sporen van elke stam werden geïncubeerd met 5 μg gezuiverd mRFP en vervolgens werden de monsters gecentrifugeerd. Sporen in de pelletfracties werden gebruikt om oppervlakte-eiwitten te extraheren die vervolgens werden geanalyseerd met behulp van Western blot (A), terwijl de supernatanten serieel werden verdund en geanalyseerd met dot blot (B). Seriële verdunningen van gezuiverd mRFP werden gebruikt als standaard. Immuno-reacties in beide panelen werden uitgevoerd met anti-His antilichaam geconjugeerd met mierikswortelperoxidase.
De adsorptie-efficiëntie van sporen van de drie stammen werd ook geanalyseerd door fluorescentiemicroscopie (figuur 4A en supplementair figuur S1). Microscopie beelden werden geanalyseerd door ImageJ software (v1.48, NIH) voor het uitvoeren van kwantitatieve fluorescentie beeldanalyse en spore fluorescentie werd berekend zoals beschreven in Materialen en Methoden. De analyse van 80 sporen van elke stam gaf een gemiddelde fluorescentiewaarde per pixel van 52,43 (± 5,94), 20,81 (± 2,71), en 32,33 (± 2,97; arbitraire eenheden) voor Q97; arbitraire eenheden) voor QM B1551, PV361, en PY79, respectievelijk (Figuur 4B), die verder bewijs dat QM B1551 sporen geadsorbeerd meer mRFP dan sporen van de andere twee stammen.
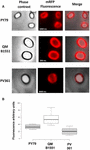
FIGUUR 4. Efficiëntie van de adsorptie van mRFP aan sporen van B. megaterium QM B1551 en PV361 en B. subtilis PY79. (A) Fluorescentiemicroscopiebeelden van PY79, QM B1551, en PV361 sporen na mRFP-adsorptie en wassen. Belichtingstijden zijn aangegeven. Fasecontrast en rode fluorescentie overlays worden getoond (samengevoegde paneel). Schaalstaven 1 um. (B) Box plots met de fluorescentie-intensiteit van tachtig verschillende sporen van elke stam. De grenzen van elk vakje vertegenwoordigen het eerste en het derde kwartiel (25 en 75%) en de waarden buiten de vakjes vertegenwoordigen de maximum- en minimumwaarden. De lijn die het hokje scheidt, geeft de mediaanwaarde voor elke stam aan. P waarde is minder dan 0.0001.
Gebaseerd op de resultaten van Figuren 3 en 4, concluderen we dat het exosporium, aanwezig in QM B1551 en ontbrekend in PV361 een relevante rol heeft in de adsorptie van mRFP.
Bovendien gaven de resultaten van figuur 4 aan dat B. subtilis PY79 sporen efficiënter zijn dan B. megaterium PV361 sporen in het adsorberen van mRFP, terwijl dot blotting gerapporteerd in figuur 3B een vergelijkbare adsorptie-efficiëntie voor de twee stammen aangaf. Wij denken dat dit verschil te wijten is aan een sterke vermindering van de fluorescentie wanneer mRFP gebonden is aan PV361 maar niet aan PY79 of QM B1551 sporen (zie hieronder).
Quantitatieve beoordeling van mRFP Adsorptie aan QM B1551 sporen
Dot blot experimenten (figuur 3B) aangegeven dat wanneer 5 ug gezuiverd mRFP werd gebruikt in adsorptiereacties met 5,0 x 108 sporen van de QM B1551 stam bijna alle heterologe eiwit was gebonden aan de spore. Om de maximale hoeveelheid mRFP te bepalen die aan QM B1551-sporen kan worden geadsorbeerd, herhaalden we de reacties met toenemende concentraties gezuiverd mRFP, d.w.z. 5,0 × 108 QM B1551-sporen werden gereageerd met 5, 10, 20, 40, 80 en 160 μg gezuiverd mRFP. Na de reacties werden sporen verzameld door centrifugatie en de supernatanten die ongebonden mRFP bevatten werden serieel verdund en geanalyseerd door dot blotting (figuur 5A). Figuur 5B toont de resultaten van een densitometrische analyse van de dot blot (??), die aangeeft dat wanneer 5-80 ug van mRFP werd gereageerd met 5 × 108 sporen, het percentage van het eiwit gebonden aan sporen was meer dan 90%. Een afname van gebonden mRFP werd waargenomen wanneer 160 pg gezuiverd eiwit werden gebruikt in de reactie. Maar zelfs wanneer 160 μg gezuiverd mRFP werd gebruikt, werd meer dan 60% van het eiwit geabsorbeerd, wat erop wijst dat 5,0 × 108 sporen van QM B1551 ongeveer 100 μg mRFP kunnen adsorberen.
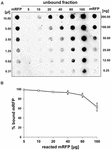
FIGUUR 5. Kwantitatieve bepaling van de adsorptie van mRFP aan B. megaterium QM B1551-sporen. 5 × 108 sporen werden geïncubeerd met 5, 10, 20, 40, 80 en 160 μg gezuiverd mRFP. De reactiemengsels werden vervolgens onderworpen aan centrifugatie en de supernatanten serieel verdund en geanalyseerd door dot blot (A). Seriële verdunningen van gezuiverd mRFP werden gebruikt als standaard. Immuno-reacties in beide panelen werden uitgevoerd met anti-His antilichaam geconjugeerd met mierikswortelperoxidase. (B) Percentage mRFP geadsorbeerd aan sporen na reactie met gedefinieerde hoeveelheden endogeen mRFP. Foutbalken tonen de standaardfouten van het gemiddelde van drie experimenten en de P-waarde nooit boven 0,0025.
mRFP Lokaliseert aan de Inter-Coat Space in B. megaterium QM B1551 Spores
Een immuno-fluorescentiemicroscopie aanpak werd gebruikt om te beoordelen of geadsorbeerde mRFP-moleculen werden blootgesteld aan het oppervlak van B. megaterium QM B1551 spores. QM B1551 sporen geadsorbeerd met verschillende hoeveelheden mRFP werden gereageerd met monoklonale anti-His antilichaam herkennen van de recombinant mRFP, vervolgens met fluorescerende anti-muis secundaire antilichaam (Santa Cruz Biotechnology, Inc) en waargenomen onder de fluorescentie microscoop (figuur 6). Met de laagste hoeveelheid mRFP gebruikt in dit experiment (2 ug) de mRFP fluorescentiesignaal (rood) werd waargenomen rondom de spore, terwijl de immunofluorescentie signaal (groen) was zwak en vooral geconcentreerd op de sporen polen, wat suggereert dat alleen in die punten mRFP werd blootgesteld op het spore-oppervlak. Naarmate de in de reactie gebruikte hoeveelheid mRFP toenam, nam het aantal groene vlekken toe (5 en 10 μg) en met de grootste hoeveelheid mRFP die in de reactie werd gebruikt (20 μg), werd een bijna volledige groene ring rond de sporen waargenomen. Op basis van de resultaten in figuur 6 hebben wij de hypothese geformuleerd dat mRFP-moleculen door het exosporium infiltreren en zich lokaliseren in de ruimte tussen de buitenlaag en het exosporium, d.w.z. dat bij gebruik van een geringe hoeveelheid mRFP bijna alle eiwitmoleculen zich intern in het exosporium bevinden en slechts op enkele plaatsen beschikbaar zijn voor het antilichaam. Toenemende hoeveelheden mRFP in adsorptiereacties resulteren in het “opvullen” van de ruimte tussen de coatings, totdat uiteindelijk meer mRFP-moleculen beschikbaar zijn voor het antilichaam op het sporeoppervlak. Deze hypothese impliceert dat als het exosporium ontbreekt, alle mRFP beschikbaar zou moeten zijn voor het antilichaam. Om dit te testen vergeleken we met behulp van immunofluorescentiemicroscopie gelijke aantallen sporen van QM B1551 (met exosporium) en van PV361 (zonder exosporium) geïncubeerd met dezelfde hoeveelheid mRFP (5 μg). Wanneer het exosporium aanwezig was (QM B1551) was mRFP slechts gedeeltelijk beschikbaar voor het antilichaam en werden groene vlekken waargenomen (figuur 7 en supplementaire figuur S2). Wanneer het exosporium niet aanwezig was (PV361) was geadsorbeerd mRFP beschikbaar voor het antilichaam rondom de spore en werd een volledige groene ring gevormd, wat de hypothese ondersteunt dat mRFP intern is aan het exosporium in QM B1551-sporen.
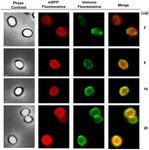
FIGUUR 6. Immunofluorescentie-analyse van mRFP-adsorbed B. megaterium QM B1551-sporen. Aliquots van 5 × 108 QM B1551 sporen werden geïncubeerd met variabele concentraties mRFP en werden vervolgens geanalyseerd door fasecontrast, fluorescentie en immunofluorescentiemicroscopie, zoals beschreven in de materialen en methodes. Hetzelfde microscopieveld voor elke reactie wordt samen met het samenvoegingspaneel gerapporteerd. De belichtingstijd was 200 ms voor alle beelden. Schaal bar, 1 um.
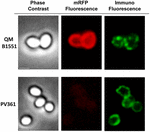
FIGUUR 7. Immunofluorescentie van mRFP, geadsorbeerd aan B. megaterium QM B1551- en PV361-sporen. 5 × 108 QM B1551 en PV361 sporen werden geïncubeerd met 5 ug mRFP en vervolgens geanalyseerd door immunofluorescentiemicroscopie, zoals beschreven in de Materials and Methods. Voor elk veld fasecontrast en immunofluorescentiemicroscopie worden getoond. De belichtingstijd was 200 ms voor alle beelden. Schaalbalk, 1 urn.
Want QM B1551 sporen gebruikt in de experimenten van figuur 7 toonde een volledige rode fluorescerende ring als in figuur 2, PV361 sporen toonde een zeer zwak rood fluorescerend signaal. Met PV361 sporen een rood signaal werd alleen waargenomen met behulp van lange belichtingstijden op de fluorescentie microscoop (figuur 4). Aangezien mRFP aanwezig is rond PV361 sporen (figuren 3 en 7), concluderen we dat mRFP fluorescentie verzwakt is wanneer het eiwit geadsorbeerd is aan PV361 sporen. Verdere experimenten zullen nodig zijn om dit punt volledig te adresseren.
Discussie
De belangrijkste bevindingen van dit rapport zijn dat sporen van B. megaterium zeer efficiënt zijn in het adsorberen van het heterologe modeleiwit mRFP, dat het exosporium een belangrijke rol speelt in dit proces, en dat mRFP-moleculen infiltreren door het exosporium en zich lokaliseren tussen de buitenste mantel en het exosporium. Deze resultaten zijn een uitbreiding van eerder werk met sporen van B. subtilis en tonen aan dat sporen van een andere soort ook kunnen gebruikt worden om heterologe eiwitten af te leveren via de adsorptiemethode. De hoge adsorptie-efficiëntie van B. megaterium-sporen is gedeeltelijk toe te schrijven aan de grote omvang van de spore in vergelijking met die van B. subtilis. De oppervlakte van de spore van B. megaterium is immers ongeveer 2,5 maal groter dan die van de spore van B. subtilis, met een oppervlakte van 5,33 μm2 (h: 1,60 ± 0,16 w: 0,84 ± 0,07) tegenover 1,98 μm2 (h: 1,07 ± 0,09 w: 0,48 ± 0,03). De grote afmetingen maken de adsorptie mogelijk van maximaal 100 μg mRFP wanneer 160 μg eiwit met sporen reageert.
De waarneming dat mRFP het exosporium passeert, wijst erop dat het doorlaatbaar is voor mRFP, een 27 kDa-eiwit. Permeabiliteit van het exosporium is niet geheel verrassend aangezien kiemstoffen aanwezig in de omgeving de externe lagen van de spore moeten doorkruisen om hun receptoren te bereiken, hoewel kiemstoffen typisch kleine moleculen zijn met moleculaire massa’s typisch <200 Da. Bovendien komen de mRFP-gegevens in grote lijnen overeen met de resultaten van eerdere studies die werden uitgevoerd met gelabelde dextrans en verwante moleculen (Koshikawa et al., 1984; Nishihara et al., 1989). In die studies werd gesuggereerd dat het exosporium van B. megaterium QM B1551 een permeabiliteitsbarrière vormt voor moleculen met een molecuulgewicht van meer dan 100 KDa, terwijl het de passage beïnvloedt van moleculen met een massa tussen 2 en 40 kDa (Koshikawa et al, 1984; Nishihara et al., 1989).
Een interessante uitdaging voor toekomstig werk zal zijn om het mechanisme of de route van infiltratie die mRFP, en bij gevolgtrekking andere heterologe eiwitten van belang, neemt om de inter-coat ruimte te voeren vast te stellen. Onderzoek met transmissie-elektronenmicroscopie van doorgesneden B. megaterium QM B1551 sporen toont aan dat het exosporium uit twee identieke “schillen” bestaat (Manetsberger et al., 2015a), en het zou kunnen dat het raakvlak tussen elk van deze structuren (in vroege publicaties beschreven als “apicale openingen”) het binnendringen van relatief grote moleculen mogelijk maakt. Het achterhalen van de basis voor het schijnbare verlies van mRFP-fluorescentie bij adsorptie aan PV361-sporen, en of mRFP-moleculen in staat zijn om de buitenste mantellagen te infiltreren, zoals waargenomen voor B. subtilis-sporen (Donadio et al., 2016), zal ook van belang zijn.
In de huidige studie, stellen we de hypothese dat mRFP-moleculen bij voorkeur het exosporium oversteken en zich ophopen in de tussenlaagruimte tussen de buitenste laag en het exosporium. In dit model worden mRFP-moleculen pas geadsorbeerd en afgebeeld op het spore-oppervlak wanneer adsorptieplaatsen (of volumetrische capaciteit?) in de ruimte tussen de lagen voldoende bezet zijn. Dit impliceert dat de adsorptiebenadering van oppervlaktedisplay kan worden gebruikt met B. megaterium QM B1551-sporen, hoewel het systeem afhankelijk is van de spore-eiwitverhouding die wordt gebruikt in adsorptiereacties. Aangezien verschillende stammen van B. megaterium reeds lang industrieel worden gebruikt voor de productie van enzymen zoals amylasen en dehydrogenasen, vitaminen en antimicrobiële moleculen (Vary et al, 2007), suggereren onze gegevens een nieuwe biotechnologische toepassing voor de spore van B. megaterium als vehikel om heterologe eiwitten te binden en af te leveren.
Author Contributions
ML en GD – voerden het grootste deel van de experimenten; RA en AS – bijgedragen aan de interpretatie van gegevens voor het werk; GiC, LB, en GrC – bijgedragen aan het opstellen van het werk en kritisch te herzien; ER – bijgedragen aan discussies en suggesties tijdens het werk en bijgedragen aan het schrijven van het manuscript; RI – leidde het werk en bijgedragen aan het schrijven van het manuscript. Alle auteurs hebben het uiteindelijke manuscript gelezen en goedgekeurd.
Funding
Dit werk werd ondersteund door een subsidie (STAR Linea1- 2014) van de Federico II Universiteit van Napels en Compagnia di San Paolo, Italië aan RI.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Supplementary Material
Het Supplementary Material voor dit artikel is online te vinden op: https://www.frontiersin.org/article/10.3389/fmicb.2016.01752/full#supplementary-material
FIGUUR S1 | Whole field beelden van fluorescentie microscopie analyse gerapporteerd in figuur 4. Dezelfde microscopie veld werd waargenomen door fasecontrast en fluorescentie microscopie. Schaal bar 5 pm. De samenvoeging panel wordt gerapporteerd. De belichtingstijd is aangegeven.
FIGUUR S2 | Hele veld beelden van Immunofluorescentie-analyse gerapporteerd in figuur 7. Dezelfde microscopie veld werd waargenomen door fasecontrast en fluorescentie microscopie. Schaal bar 5 pm. De belichtingstijd was 200 ms voor alle beelden.
TABLE S1 | Densitometrische analyse van dot blot experimenten met de supernatanten van de adsorptiereactie met QM B1551 sporen (figuur 1B).
TABLE S2 | Densitometrische analyse van dot blot experimenten met de supernatanten van verschillende wasbeurten na de adsorptiereactie (figuur 1C).
TABLE S3 | Densitometrische analyse van dot blot experimenten met de supernatanten van de adsorptiereactie uitgevoerd met sporen van stammen PY79, QM B1551, en PV361 (figuur 3B).
TABLE S4 | Densitometrische analyse van dot blot experimenten met de supernatanten van de adsorptiereactie uitgevoerd met verschillende hoeveelheden mRFP.
Baccigalupi, L., Ricca, E., and Ghelardi, E. (2015). “Non-LAB probiotics: spore Formers,” in Probiotics and Prebiotics: Current Research and Future Trends, eds K. Venema and A. P. do Carmo (Norfolk, VA: Caister Academic Press), 93-103. doi: 10.21775/9781910190098.06
CrossRef Full Text | Google Scholar
Campbell, R. E., Tour, O., Palmer, A. E., Steinbach, P. A., Baird, G. S., Zacharias, D. A., et al. (2002). Een monomeer rood fluorescerend eiwit. Proc. Natl. Acad. Sci. U.S.A. 99, 7877-7882. doi: 10.1073/pnas.082243699
PubMed Abstract | CrossRef Full Text | Google Scholar
Cangiano, G., Sirec, T., Panarella, C., Isticato, R., Baccigalupi, L., De Felice, M., et al. (2014). De sps genproducten beïnvloeden de kieming, hydrofobiciteit en eiwitadsorptie van Bacillus subtilis sporen. Appl. Environ. Microbiol. 80, 7293-7302. doi: 10.1128/AEM.02893-14
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutting, S. M. (2011). Bacillus probiotica. Food Microbiol. 28, 214-220. doi: 10.1016/j.fm.2010.03.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutting, S., and Vander Horn, P. B. (1990). “Genetic analysis,” in Molecular Biological Methods for Bacillus, eds C. Harwood and S. Cutting (Chichester: John Wiley and Sons), 27-74.
Google Scholar
Díaz-González, F., Milano, M., Olguin-Araneda, V., Pizarro-Cerda, J., Castro-Córdova, P., Tzeng, S. C., et al. (2015). Eiwitsamenstelling van de buitenste exosporium-achtige laag van Clostridium difficile 630 sporen. J. Proteom. 123, 1-13. doi: 10.1016/j.jprot.2015.03.035
PubMed Abstract | CrossRef Full Text | Google Scholar
Di Luccia, B., D’Apuzzo, E., Varriale, F., Baccigalupi, L., Ricca, E., and Pollice, A. (2016). Bacillus megaterium SF185 induceert stress pathways en beïnvloedt de celcyclusverdeling van humane darmepitheelcellen. Benef. Microb. 7, 609-620. doi: 10.3920/BM2016.0020
PubMed Abstract | CrossRef Full Text | Google Scholar
Donadio, G., Lanzilli, M., Sirec, T., Ricca, E., and Isticato, R. (2016) Eiwitadsorptie en -lokalisatie op wild type en mutant sporen van Bacillus subtilis. Microb. Cell Fact. 15:153. doi: 10.1186/s12934-016-0551-2
PubMed Abstract | CrossRef Full Text
Eppinger, M., Bunk, B., Johns, M. A., Edirisinghe, J. N., Kutumbaka, K. K., Koenig, S. S., et al. (2011). Genoomsequenties van de biotechnologisch belangrijke Bacillus megaterium stammen QM B1551 en DSM319. J. Bacteriol. 193, 4199-4213. doi: 10.1128/JB.00449-11
PubMed Abstract | CrossRef Full Text | Google Scholar
Giglio, R., Fani, R., Isticato, R., De Felice, M., Ricca, E., and Baccigalupi, L. (2011). Organisatie en evolutie van de cotG- en cotH-genen van Bacillus subtilis. J. Bacteriol. 193, 6664-6673. doi: 10.1128/jb.06121-11
PubMed Abstract | CrossRef Full Text | Google Scholar
Harwood, C. R., and Cutting, S. M. (1990). Molecular Biological Methods for Bacillus. Chichester: John Wiley and Sons.
Google Scholar
Huang, J. M., Hong, H. A., Van Tong, H., Hoang, T. H., Brisson, A., and Cutting, S. M. (2010). Mucosale levering van antigenen met behulp van adsorptie aan bacteriële sporen. Vaccine 28, 1021-1030. doi: 10.1016/j.vaccine.2009.10.127
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., Pelosi, A., De Felice, M., and Ricca, E. (2010). CotE bindt aan CotC en CotU en bemiddelt hun interactie tijdens de vorming van de sporenlaag in Bacillus subtilis. J. Bacteriol. 192, 949-954. doi: 10.1128/JB.01408-09
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., Pelosi, A., Zilhão, R., Baccigalupi, L., Henriques, A. O., De Felice, M., et al. (2008). CotC-CotU heterodimerisatie tijdens de assemblage van de spore coat van Bacillus subtilis. J. Bacteriol. 190, 1267-1275. doi: 10.1128/JB.01425-07
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., and Ricca, E. (2014). Spore oppervlakte weergave. Microbiol. Spectr. 2, 351-366. doi: 10.1128/microbiolspec.TBS-0011-2012
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., Sirec, T., Treppiccione, L., Maurano, F., De Felice, M., Rossi, M., et al. (2013). Nonrecombinante weergave van de B-subeenheid van het hitte labiele toxine van Escherichia coli op wild type en mutante sporen van Bacillus subtilis. Microb. Cell Fact. 12:98. doi: 10.1186/1475-2859-12-98
PubMed Abstract | CrossRef Full Text | Google Scholar
Knecht, L. D., Pasini, P., and Daunert, S. (2011). Bacteriële sporen als platforms voor bioanalytische en biomedische toepassingen. Anal. Bioanal. Chem. 400, 977-989. doi: 10.1007/s00216-011-4835-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Koshikawa, T., Beaman, T. C., Pankratz, H. S., Nakashio, S., Corner, T. R., and Gerhardt, P. (1984). Resistance, germination, and permeability correlates of Bacillus megaterium spores successively divested of integument layers. J. Bacteriol. 159, 624-632.
PubMed Abstract | Google Scholar
Manetsberger, J., Hall, E. A. H., and Christie, G. (2015a). Plasmide-gecodeerde genen beïnvloeden exosporium assemblage en morfologie in Bacillus megaterium QM B1551 sporen. FEMS Microbiol. Lett. 362:fnv147. doi: 10.1093/femsle/fnv147
PubMed Abstract | CrossRef Full Text | Google Scholar
Manetsberger, J., Manton, J. D., Erdelyi, M. J., Lin, H., Rees, D., Christie, G., et al. (2015b). Ellipsoïde lokalisatie microscopie infereert de grootte en volgorde van eiwitlagen in Bacillus spore coat. Biophys. J. 109, 2058-2066. doi: 10.1016/j.bpj.2015.09.023
PubMed Abstract | CrossRef Full Text | Google Scholar
McCloy, R., Rogers, S., Caldon, E., Lorca, T., Castro, A., and Burgess, A. (2014). Gedeeltelijke remming van Cdk1 in G2 fase overruled de SAC en ontkoppelt mitotische gebeurtenissen. Cell Cycle 13, 1400-1412. doi: 10.4161/cc.28401
PubMed Abstract | CrossRef Full Text | Google Scholar
McKenney, P. T., Driks, A., and Eichemberger, P. (2012). De Bacillus subtilis endospore: assemblage en functies van de meerlagige mantel. Nat. Rev. Microbiol. 11, 33-44. doi: 10.1038/nrmicro2921
PubMed Abstract | CrossRef Full Text | Google Scholar
Nicholson, W. L., and Setlow, P. (1990). “Sporulation, germination and out-growth,” in Molecular biological methods for Bacillus, eds C. Harwood and S. Cutting (Chichester: John Wiley and Sons), 391-450.
Google Scholar
Nishihara, T., Takubo, Y., Kawamata, E., Koshikawa, T., Ogaki, J., and Kondo, M. (1989). Rol van de buitenmantel in de resistentie van Bacillus megaterium spore. J. Biochem. 106, 270-273.
PubMed Abstract | Google Scholar
Pesce, G., Rusciano, G., Sirec, T., Isticato, R., Sasso, A., and Ricca, E. (2014). Metingen van oppervlaktelading en hydrodynamische coëfficiënt van Bacillus subtilis spore met behulp van een optisch pincet. Colloids Surf. B Biointerfaces 116C, 568-575. doi: 10.1016/j.colsurfb.2014.01.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Ricca, E., Baccigalupi, L., Cangiano, G., De Felice, M., and Isticato, R. (2014). Mucosale vaccintoediening door niet-recombinante sporen van Bacillus subtilis. Microb. Cell Fact. 13, 115. doi: 10.1186/s12934-014-0115-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosso, M. L., and Vary, P. S. (2005). Distributie van Bacillus megaterium QM B1551 plasmiden onder andere B. megaterium stammen en Bacillus species. Plasmid 53, 205-217. doi: 10.1016/j.plasmid.2004.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Rusciano, G. Zito, G., Isticato, R., Sirec, T., Ricca, E., Bailo, E., and Sasso, A. (2014). Nanoschaal chemische beeldvorming van Bacillus subtilis sporen door het combineren van tip-enhanced Raman scattering en geavanceerde statistische tools. ACS Nano 8, 12300-12309. doi: 10.1021/nn504595k
PubMed Abstract | CrossRef Full Text | Google Scholar
Sirec, T., Strazzulli, A., Isticato, R., De Felice, M., Moracci, M., and Ricca, E. (2012). Adsorptie van beta-galactosidase van Alicyclobacillus acidocaldarius aan wild type en mutanten sporen van Bacillus subtilis. Microb. Cell Fact. 11:100. doi: 10.1186/1475-2859-11-100
PubMed Abstract | CrossRef Full Text | Google Scholar
Stewart, G. C. (2015). De Exosporium laag van bacteriële sporen: een verbinding met de omgeving en de geïnfecteerde gastheer. Microbiol. Mol. Biol. Rev. 79, 437-457. doi: 10.1128/MMBR.00050-15
PubMed Abstract | CrossRef Full Text | Google Scholar
Vary, P. S., Biedendieck, R., Fuerch, T., Meinhardt, F., Rohde, M., Deckwer, W. -D., et al. (2007). Bacillus megaterium – van eenvoudige bodembacterie tot gastheer voor industriële eiwitproductie. Toepassing Microbiol. Biotechnol. 76, 957-967 doi: 10.1007/s00253-007-1089-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Youngman, P., Perkins, J. B., and Losick, R. (1984). A novel method for the rapid cloning in Escherichia coli of Bacillus subtilis chromosomal DNA adjacent to Tn917 insertion. Mol. Gen. Genet. 195, 424-433. doi: 10.1007/BF00341443
CrossRef Full Text | Google Scholar