Uwalnianie hormonówEdit
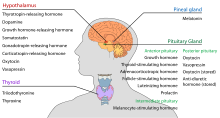
Podwzgórze pełni centralną funkcję neuroendokrynną, przede wszystkim poprzez kontrolę przedniej części przysadki, która z kolei reguluje różne gruczoły dokrewne i narządy. Hormony uwalniające (zwane również czynnikami uwalniającymi) są produkowane w jądrach podwzgórza, a następnie transportowane wzdłuż aksonów do środkowego zwoju lub tylnej części przysadki, gdzie są przechowywane i uwalniane w razie potrzeby.
Przysadka przednia
W osi podwzgórze-adenohypophyseal, hormony uwalniające, znane również jako hormony hipofizjotropowe lub hormony podwzgórza, są uwalniane z pośrodkowej eminencji, przedłużenie podwzgórza, do układu wrotnego podwzgórza, który przenosi je do przedniej przysadki, gdzie wywierają swoje funkcje regulacyjne na wydzielanie hormonów adenohypophyseal. Hormony hipofizjotropowe są stymulowane przez komórki neurosekrecyjne zlokalizowane w okolicy okołokomorowej podwzgórza. Po uwolnieniu do naczyń włosowatych komory trzeciej, hormony podwzgórzowe przemieszczają się przez tzw. krążenie wrotne podwzgórzowo-przysadkowe. Po dotarciu do miejsca przeznaczenia w przedniej części przysadki, hormony te wiążą się ze specyficznymi receptorami znajdującymi się na powierzchni komórek przysadki. W zależności od tego, które komórki są aktywowane przez to wiązanie, przysadka zacznie wydzielać lub przestanie wydzielać hormony do reszty krwiobiegu.
| Wydzielany hormon | Skrót | Wydzielany przez | Skutek |
|---|---|---|---|
| Hormon uwalniający tyreotropinę (Prolactin-releasing hormone) |
TRH, TRF, lub PRH | Parvocellular neurosecretory cells of the paraventricular nucleus | Stimulate thyroid-stimulating hormone (TSH) release from anterior pituitary (primarily) Stimulate prolactin release from anterior pituitary |
| Corticotropin-uwalniająca hormon | CRH lub CRF | Komórki neurosekrecyjne jądra przykomorowego | Stymulują uwalnianie hormonu adrenokortykotropowego (ACTH) z przedniej części przysadki |
| Dopamina (Prolaktyna-hamujący hormon prolaktyny) |
DA lub PIH | Neurony dopaminergiczne jądra łukowatego | Zahamowanie uwalniania prolaktyny z przedniej części przysadki |
| Hormon wzrostu-hormon uwalniający hormon wzrostu | GHRH | Neuroendokrynne neurony jądra łukowatego | Stymulują uwalnianie hormonu wzrostu (GH) z przedniej części przysadki.hormonu wzrostu (GH) z przedniej części przysadki |
| Hormon uwalniający gonadotropiny | GnRH lub LHRH | Komórki neuroendokrynne okolicy przedwzgórza | Stymulują wydzielanie hormonu folikularnego (FSH).stymulujące uwalnianie hormonu folikularnego (FSH) z przedniej części przysadki Stymulują uwalnianie hormonu luteinizującego (LH) z przedniej części przysadki |
| Somatostatyna (hormon hamujący wzrost) |
SS, GHIH, lub SRIF | Komórki neuroendokrynne jądra okołokomorowego | Zahamowanie uwalniania hormonu wzrostu (GH) z przedniej części przysadki Zahamowanie (umiarkowane) uwalniania hormonu tyreotropowego (TSH) z przedniej części przysadki |
Inne hormony wydzielane z eminencji przyśrodkowej obejmują wazopresynę, oksytocynę i neurotensynę.
Przysadka tylna
W osi podwzgórze-jądro, hormony neurofizjologiczne są uwalniane z przysadki tylnej, która jest właściwie przedłużeniem podwzgórza, do krążenia.
| Hormon wydzielany | Skrót | Wytwarzany przez | Działanie |
|---|---|---|---|
| Oksytocyna | OXY lub OXT | Magnokomórkowe komórki neurosekrecyjne jądra przykomorowego i jądra nadocznego Jądro | Skurcz macicy Laktacja (odruch wypróżniania) |
| Wazopresyna (hormon antydiuretyczny) |
ADH lub AVP | Magnokomórkowe i parwokomórkowe komórki neurosekrecyjne jądra przyśrodkowego, komórki magnokomórkowe w jądrze nadwzrokowym | Zwiększenie przepuszczalności dla wody komórek kanalika dystalnego i przewodu zbiorczego w nerce, co umożliwia wchłanianie zwrotne wody i wydalanie zagęszczonego moczu |
Wiadomo również, że hormony osi podwzgórze-przysadka-nadnercza (HPA) są związane z niektórymi chorobami skóry i homeostazą skóry. Istnieją dowody łączące nadczynność hormonów HPA z chorobami skóry związanymi ze stresem i nowotworami skóry.
StymulacjaEdit
Podwzgórze koordynuje wiele hormonalnych i behawioralnych rytmów okołodobowych, złożonych wzorów wyjść neuroendokrynnych, złożonych mechanizmów homeostatycznych i ważnych zachowań. Podwzgórze musi zatem reagować na wiele różnych sygnałów, z których niektóre są generowane zewnętrznie, a niektóre wewnętrznie. Sygnalizacja fali delta powstająca we wzgórzu lub w korze wpływa na wydzielanie hormonów uwalniających; GHRH i prolaktyna są stymulowane, podczas gdy TRH jest hamowana.
Podwzgórze jest wrażliwe na:
- Światło: długość dnia i fotoperiod do regulacji rytmów okołodobowych i sezonowych
- Bodźce węchowe, w tym feromony
- Steroidy, w tym steroidy gonadalne i kortykosteroidy
- Informacje przekazywane drogą nerwową pochodzące w szczególności z serca, jelitowego układu nerwowego (przewodu pokarmowego) i układu rozrodczego.
- Wejścia autonomiczne
- Bodźce płynące z krwi, w tym leptyna, grelina, angiotensyna, insulina, hormony przysadki mózgowej, cytokiny, stężenie glukozy w osoczu i osmolarność itp.
- Stres
- Inwazja mikroorganizmów poprzez zwiększenie temperatury ciała, resetowanie termostatu organizmu w górę.
Budziki węchoweEdit
Budziki węchowe są ważne dla reprodukcji seksualnej i funkcji neuroendokrynnych u wielu gatunków. Na przykład, jeśli ciężarna mysz jest narażona na mocz „obcego” samca w krytycznym okresie po współżyciu, to ciąża się nie udaje (efekt Bruce’a). Tak więc, podczas stosunku samica myszy tworzy precyzyjną „pamięć węchową” swojego partnera, która utrzymuje się przez kilka dni. Pheromonal wskazówki pomocy synchronizacji rui w wielu gatunków; u kobiet, synchronizowane miesiączki mogą również wynikać z feromonalne wskazówki, chociaż rola feromonów u ludzi jest disputed.
Blood-borne stimuliEdit
Peptide hormony mają ważne wpływy na podwzgórze, i aby to zrobić muszą przejść przez barierę krew-mózg. Podwzgórze jest ograniczone częściowo przez wyspecjalizowane regiony mózgu, które nie mają skutecznej bariery krew-mózg; śródbłonek kapilar w tych miejscach jest fenestrowany, aby umożliwić swobodne przejście nawet dużych białek i innych cząsteczek. Niektóre z tych miejsc są miejscami neurosekrecji – neurohypophysis i eminencja przyśrodkowa. Inne jednak są miejscami, w których mózg pobiera próbki składu krwi. Dwa z tych miejsc, SFO (subfornical organ) i OVLT (organum vasculosum of the lamina terminalis) są tzw. narządami okrężnymi, w których neurony pozostają w ścisłym kontakcie zarówno z krwią, jak i płynem mózgowo-rdzeniowym. Struktury te są gęsto unaczynione i zawierają neurony osmoreceptorowe i sodoreceptorowe, które kontrolują picie, uwalnianie wazopresyny, wydalanie sodu i apetyt na sód. Zawierają one również neurony z receptorami dla angiotensyny, przedsionkowego czynnika natriuretycznego, endoteliny i relaksyny, z których każdy jest ważny w regulacji równowagi płynów i elektrolitów. Neurony w OVLT i SFO rzutują się na jądro nadwzrokowe i jądro przyśrodkowe, a także na obszary podwzgórza przedwzrokowego. Narządy okołokomorowe mogą być również miejscem działania interleukin w celu wywołania zarówno gorączki, jak i wydzielania ACTH, poprzez wpływ na neurony przyśrodkowe.
Nie jest jasne, jak wszystkie peptydy, które wpływają na aktywność podwzgórza, uzyskują niezbędny dostęp. W przypadku prolaktyny i leptyny istnieją dowody na aktywny wychwyt w splocie naczyniówkowym z krwi do płynu mózgowo-rdzeniowego (CSF). Niektóre hormony przysadki mózgowej mają ujemne sprzężenie zwrotne na wydzielanie podwzgórzowe; na przykład hormon wzrostu działa z powrotem na podwzgórze, ale w jaki sposób dostaje się do mózgu nie jest jasne. Istnieje również dowód na centralne działania prolactin.
Wyniki sugerują, że hormon tarczycy (T4) jest podejmowane przez komórki glejowe podwzgórza w infundibular nucleus / median eminence, i że jest tutaj przekształcony w T3 przez typ 2 dejodynazy (D2). Następnie T3 jest transportowana do neuronów produkujących hormon uwalniający tyreotropinę (TRH) w jądrze przyśrodkowym. W neuronach tych znaleziono receptory dla hormonów tarczycy, co wskazuje, że są one rzeczywiście wrażliwe na bodźce T3. Ponadto, neurony te wykazują ekspresję MCT8, transportera hormonów tarczycy, co potwierdza teorię, że T3 jest do nich transportowany. T3 może następnie związać się z receptorem hormonu tarczycy w tych neuronach i wpływać na produkcję hormonu uwalniającego tyreotropinę, regulując w ten sposób produkcję hormonów tarczycy.
Podwzgórze funkcjonuje jako rodzaj termostatu dla ciała. Ustawia pożądaną temperaturę ciała i stymuluje albo produkcji ciepła i retencji, aby podnieść temperaturę krwi do wyższego ustawienia lub pocenie się i rozszerzenie naczyń krwionośnych do chłodzenia krwi do niższej temperatury. Wszystkie gorączki wynikają z podwyższonego ustawienia w podwzgórzu; podwyższona temperatura ciała z jakiejkolwiek innej przyczyny jest klasyfikowana jako hipertermia. Rzadko, bezpośrednie uszkodzenie podwzgórza, np. w wyniku udaru, powoduje gorączkę; jest to czasami nazywane gorączką podwzgórzową. Jednak częściej takie uszkodzenie powoduje nieprawidłowo niską temperaturę ciała.
SteroidyEdit
Podwzgórze zawiera neurony, które silnie reagują na steroidy i glikokortykoidy – (hormony steroidowe nadnerczy, uwalniane w odpowiedzi na ACTH). Zawiera również wyspecjalizowane neurony wrażliwe na glukozę (w jądrze łukowatym i brzuszno-przyśrodkowym podwzgórza), które są ważne dla apetytu. Obszar preoptyczny zawiera neurony termowrażliwe; są one ważne dla wydzielania TRH.
NeuralEdit
Wydzielanie oksytocyny w odpowiedzi na ssanie lub stymulację pochwowo-szyjkową jest mediowane przez niektóre z tych szlaków; wydzielanie wazopresyny w odpowiedzi na bodźce sercowo-naczyniowe pochodzące z chemoreceptorów w ciele szyjnym i łuku aorty oraz z receptorów objętościowych przedsionka o niskim ciśnieniu jest mediowane przez inne. U szczura stymulacja pochwy również powoduje wydzielanie prolaktyny, co skutkuje pseudo ciążą po niepłodnym kryciu. U królika, współżycie wywołuje odruchową owulację. U owcy, stymulacja szyjki macicy w obecności wysokich poziomów estrogenów może wywołać zachowania macierzyńskie u dziewiczej owcy. Wszystkie te efekty pośredniczone są przez podwzgórze, a informacje przenoszone są głównie drogami rdzeniowymi, które znajdują się w pniu mózgu. Stymulacja brodawek sutkowych stymuluje uwalnianie oksytocyny i prolaktyny oraz hamuje uwalnianie LH i FSH.
Bodźce sercowo-naczyniowe są przenoszone przez nerw błędny. Pochwa przekazuje również różne informacje trzewne, w tym na przykład sygnały wynikające z rozciągnięcia żołądka lub opróżniania, w celu tłumienia lub promowania karmienia, sygnalizując uwalnianie leptyny lub gastryny, odpowiednio. Ponownie te informacje docierają do podwzgórza poprzez przekaźniki w pniu mózgu.
Dodatkowo funkcja podwzgórza odpowiada na-i regulowane przez-poziomy wszystkich trzech klasycznych neuroprzekaźników monoaminowych, noradrenaliny, dopaminy i serotoniny (5-hydroksytryptaminy), w tych drogach, z których otrzymuje unerwienie. Na przykład, noradrenergiczne wejścia powstające z locus coeruleus mają ważny wpływ regulacyjny na poziom hormonu uwalniającego kortykotropinę (CRH).
Kontrola przyjmowania pokarmuEdit
| Peptydy zwiększające zachowania żywieniowe |
. behavior | Peptides that decrease feeding behavior |
|---|---|---|
| Ghrelin | Leptin | |
| Neuropeptyd Y | (α,β,γ)-Hormony stymulujące melanocyty | |
| Peptyd związany z agouti | Peptydy transkryptowe regulowane kokainą i amfetaminą | |
| Oreksyny (A,B) | Hormon uwalniający kortykotropinę | |
| Hormon koncentrujący melaninę | Cholecystokinina | |
| Galanina | Insulina | |
| Glukagon-like peptide 1 |
Skrajnie boczna część jądra brzuszno-przyśrodkowego podwzgórza jest odpowiedzialna za kontrolę przyjmowania pokarmu. Stymulacja tego obszaru powoduje zwiększone spożycie pokarmu. Obustronne uszkodzenie tego obszaru powoduje całkowite zaprzestanie przyjmowania pokarmu. Przyśrodkowe części jądra mają kontrolujący wpływ na część boczną. Obustronne uszkodzenie przyśrodkowej części jądra brzuszno-przyśrodkowego powoduje hiperfagię i otyłość zwierzęcia. Dalsze uszkodzenie bocznej części jądra brzusznego u tego samego zwierzęcia powoduje całkowite zaprzestanie przyjmowania pokarmu.
Istnieją różne hipotezy związane z tą regulacją:
- Lipostatyczna hipoteza: Hipoteza ta utrzymuje, że tkanka tłuszczowa produkuje sygnał humoralny, który jest proporcjonalny do ilości tłuszczu i działa na podwzgórze, aby zmniejszyć spożycie żywności i zwiększyć produkcję energii. Zostało udowodnione, że hormon leptyny działa na podwzgórze, aby zmniejszyć spożycie żywności i zwiększenie produkcji energii.
- Gutpeptide hipoteza: hormony przewodu pokarmowego jak Grp, glukagony, CCK i inne twierdził, że hamują spożycie żywności. Jedzenie wchodzące do przewodu pokarmowego wyzwala uwalnianie tych hormonów, które działają na mózg w celu wytworzenia sytości. Mózg zawiera zarówno CCK-A i CCK-B receptorów.
- Glucostatic hipotezy: Aktywność ośrodka sytości w jądrach przykomorowych jest prawdopodobnie regulowana przez wykorzystanie glukozy w neuronach. Postuluje się, że gdy ich wykorzystanie glukozy jest niskie i w konsekwencji, gdy różnica glukozy w tętniczo-żylnej krwi jest niska, aktywność neuronów maleje. W tych warunkach aktywność ośrodka żerowania jest niekontrolowana, a osoba odczuwa głód. Pobór żywności jest szybko zwiększona przez dokomorowego podawania 2-deoksyglukozy zatem zmniejszenie wykorzystania glukozy w cells.
- Thermostatic hipotezy: Zgodnie z tą hipotezą, spadek temperatury ciała poniżej danej wartości zadanej stymuluje apetyt, podczas gdy wzrost powyżej wartości zadanej hamuje apetyt.
Przetwarzanie strachuEdit
Przyśrodkowa strefa podwzgórza jest częścią obwodu, który kontroluje motywowane zachowania, takie jak zachowania obronne. Analizy znakowania Fos wykazały, że seria jąder w „kolumnie kontroli zachowania” jest ważna w regulacji ekspresji wrodzonych i uwarunkowanych zachowań obronnych.
Antypredacyjne zachowania obronne
Ekspozycja na drapieżnika (takiego jak kot) wywołuje zachowania obronne u gryzoni laboratoryjnych, nawet jeśli zwierzę nigdy nie było narażone na kontakt z kotem. W podwzgórzu, ekspozycja ta powoduje wzrost liczby komórek znakowanych Fos w przednim jądrze podwzgórza, grzbietowo-przyśrodkowej części jądra brzuszno-przyśrodkowego oraz w brzuszno-bocznej części jądra przedpamięciowego (PMDvl). Jądro przedruchowe odgrywa istotną rolę w ekspresji zachowań obronnych wobec drapieżnika, gdyż zmiany w tym jądrze powodują zniesienie zachowań obronnych, takich jak zamrożenie i ucieczka. PMD nie moduluje zachowań obronnych w innych sytuacjach, ponieważ uszkodzenia tego jądra miały minimalny wpływ na wynik zamrażania po szoku. PMD ma ważne połączenia z grzbietową szarością okołowodociągową, ważną strukturą w ekspresji strachu. Ponadto, zwierzęta wykazują zachowania oceniające ryzyko w stosunku do środowiska, które wcześniej kojarzyło im się z kotem. Analiza komórek znakowanych Fos wykazała, że PMDvl jest najbardziej aktywowaną strukturą w podwzgórzu, a inaktywacja muscimolem przed ekspozycją na kontekst znosi zachowania obronne. Dlatego podwzgórze, głównie PMDvl, ma ważną rolę w ekspresji wrodzonych i uwarunkowanych zachowań obronnych do drapieżnika.
Porażka społeczna
Podobnie, podwzgórze ma rolę w porażce społecznej: Jądra w strefie przyśrodkowej są również mobilizowane podczas spotkania z agresywnym kongenerem. Pokonane zwierzę wykazuje wzrost poziomu Fos w strukturach dymorficznych płciowo, takich jak przyśrodkowe jądro przedoptyczne, brzuszno-boczna część jądra brzuszno-przyśrodkowego i brzuszne jądro przedmigdałkowe. Struktury te są ważne w innych zachowaniach społecznych, takich jak zachowania seksualne i agresywne. Ponadto, jądro przedpamięciowe jest również zmobilizowane, część grzbietowo-przyśrodkowa, ale nie część brzuszno-boczna. Lesje w tym jądrze znoszą pasywne zachowania obronne, takie jak zamrożenie i postawa „na plecach”.
.