(“menneske” / Neandertal, Tyskland)

SITES
Se “Neandertal sites” kortet, figur 35.2
PERSONER
Tå mange at nævne (se teksten for nogle fremtrædende forskere)
INDLEDNING
En af de bedst kendte og mest gådefulde af de arkaiske homininer var neandertalerne, Homo neanderthalensis. Selv om Homo neanderthalensis oprindeligt blev inkluderet i vores egen slægt og art, men adskilt med status som underart, dvs. Homo sapiens neanderthalensis, tyder stadig flere beviser fra DNA-analyser på, at de to slægter har delt sig engang før 300 kya og, hvis de nye DNA-beviser er korrekte, muligvis før 800 kya. DNA-beviserne viser imidlertid, at de har krydset hinanden, muligvis da AMH udvandrede fra Afrika en eller flere gange eller levede sammen med neandertalerne i Mellemøsten. Eurasiere og australasiater bærer i gennemsnit 2,5 % neandertalgener. Så selv om RAO-modellen for AMH’s oprindelse stadig foretrækkes, og neandertalerne betragtes som en separat art, var i det mindste nogle populationer i stand til at krydses med hinanden og var således ikke egentlige biologiske arter på det tidspunkt og i det geografiske rum.
Det materiale, der blev holotypen for arten, blev fundet i Neander-dalen nær Düsseldorf, Tyskland. Det tyske ord for dal er “thal”, og “h’et” er stumt. “h” er blevet udeladt for det almindelige navn i nogle kilder. Jeg kan kun forestille mig, at nogen blev trætte af, at folk udtaler “th” og besluttede at stå i spidsen for en kampagne for at sætte en stopper for det!
PHYLOGENY
Der er to mulige scenarier for oprindelsen af neandertalerne og AMH. Det første er baseret på fossile beviser og det andet på DNA. Fortolkningen af de fossile fund tyder på, at begge arter stammer fra H. heidelbergensis, som igen sandsynligvis har udviklet sig fra en afledt form af H. ergaster (muligvis H. mauritanicus) i Afrika. På et tidspunkt før 500 kya delte H. heidelbergensis sig i AMH- og neandertal-linjerne.
Den seneste genetiske dokumentation understøtter en opsplitning mellem AMH- og neandertal-linjerne før 800 kya i Afrika. Den gren, der fører til neandertalerne og Denisovans, betragtes derefter som H. heidelbergensis. Denisovanerne og neandertalerne deler sig derefter ~640 kya. Lokaliserede grupper fortsatte dog med at krydses med hinanden.
Det er forfriskende at lære, at populationer af homininer har krydset med hinanden og opretholdt eller dannet genetiske relationer siden begyndelsen af “vores” tid. Vi moderne mennesker er meget tættere beslægtede med hinanden end disse gamle hominin-“arter” var, og alligevel kan nogle af os ikke se os selv i andre på grund af fysiske forskelle, der ikke betyder andet end at vi gik forskellige veje på forskellige tidspunkter og tilpassede os forskellige miljøer.
Uafhængigt af neandertal/human/Denisovan-fylogenien flyttede en gruppe af H. heidelbergensis ind i Vesteuropa, hvor en lokaliseret gruppe derefter udviklede sig til neandertal-linjen <300 kya. Overgangsformer kan ses på flere lokaliteter i Vesteuropa, især i Spanien, Frankrig og Tyskland.
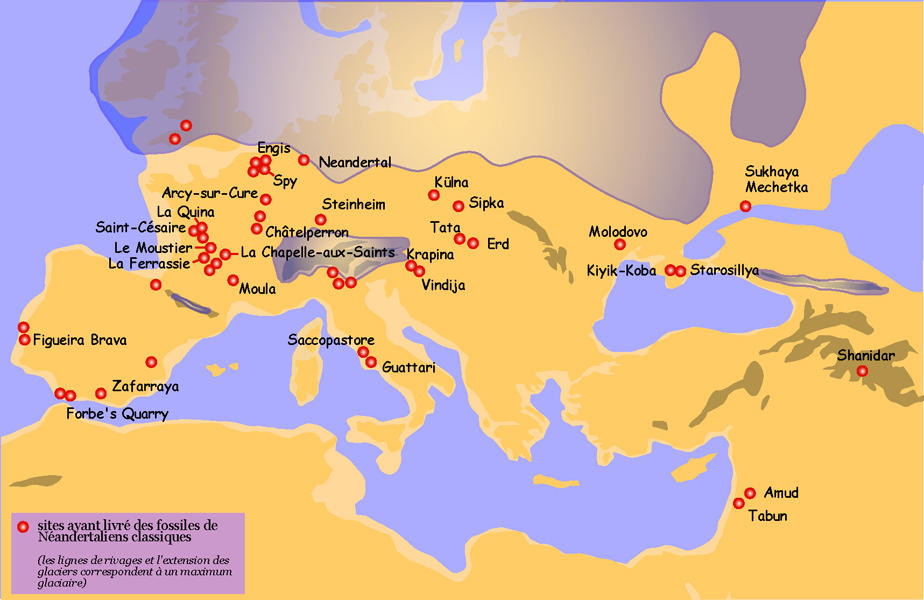
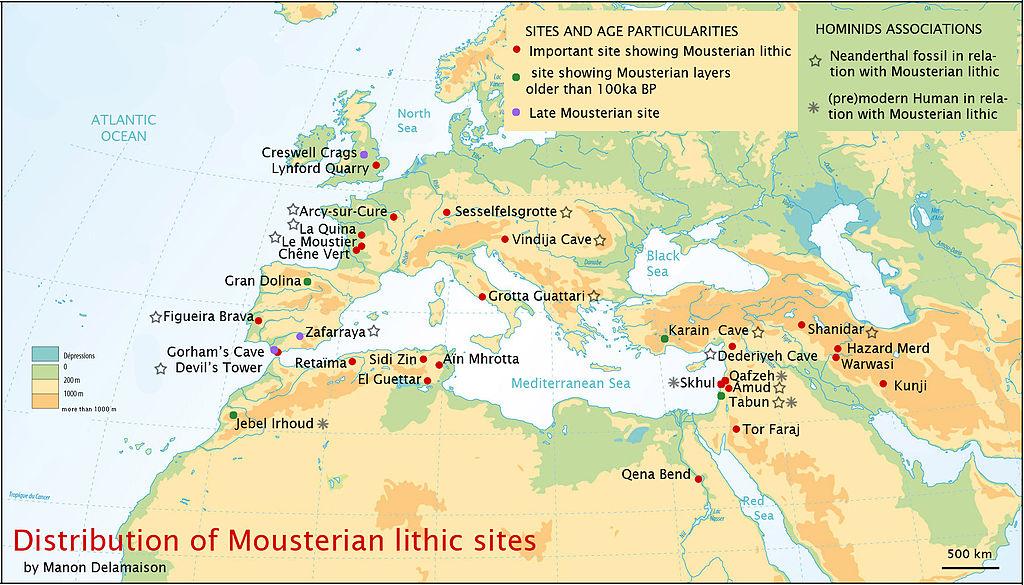
Da det pleistocæne Europa blev koldere, tilpassede neandertalerne sig til de hårdere forhold. Neandertalerne fra Vesteuropa, med deres forkrøblede og kuldetilpassede kroppe, er kendt som de “klassiske” neandertaler, til forskel fra neandertalerne i øst og sydøst, der bevarede en mere gracile morfologi. Datoer for de klassiske neandertaler varierer fra 75 til <30 kya. Figur 35.2 viser neandertal-lokaliteter i Eurasien.
DISCOVERY AND GEOGRAPHIC RANGE
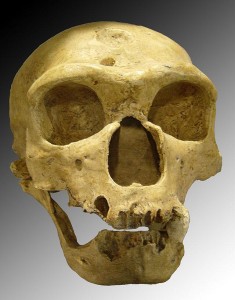
De tidligste anerkendte opdagelser blev gjort i Belgien og Gibraltar. Den næste opdagelse var resterne fra Neandertal, som gav arten sit navn. Fossilfundsteder er allestedsnærværende i Vesteuropa, hvoraf de fleste ligger i Frankrigs velvandede floddale. Mere end 200 fundsteder ligger inden for en radius af 20 miles fra Les Ezies i Frankrig. Der findes også fundsteder i Tyskland, Belgien, Spanien, Portugal og Italien. Nogle af de mere berømte steder er La Chapelle-aux-Saints, La Ferrassie og St. Cesaire i Frankrig, den føromtalte Neander-dal i Tyskland og Zafarraya-hulen i Spanien. Chapelle-aux-Saints-stedet har spillet en central rolle i udviklingen af myten om neandertalerne som klodsede, barbariske hulemænd. Resterne af en ca. 40-årig mand (se figur 35.3) blev udgravet i 1908 og analyseret af Marcellin Boule, som karakteriserede individet som primitivt, brutalt og krumrygget. Forskerne indså senere, at den voksne var ramt af gigt, hvilket forklarede hans kropsholdning. Selv om vi ikke kan vide, hvordan neandertalerne opførte sig i forhold til os selv, opnåede de et hidtil uset niveau af kulturel og teknologisk kompleksitet. Den nedsættende karakteristik holdt sig i mange år, indtil forskerne indså, hvor meget disse gamle “folkeslag” havde udrettet, såsom bevidst begravelse af deres døde.
Fra deres formodede vesteuropæiske oprindelse spredte de sig østpå til Mellemøsten og så langt østpå som Usbekistan og nordøstpå til Rusland, i Denisovanernes område. Nogle forskere accepterer ikke, at den niårige dreng på stedet Teshik Tash i Usbekistan er neandertaler, men argumenterer snarere for, at han er AMH.
Non-klassiske neandertal-lokaliteter er fundet i Kroatien, Tjekkiet, Ungarn, Syrien, Republikken Georgien, Rusland, Ukraine, Irak, Usbekistan, Ukraine, Irak og Israel. Berømte lokaliteter omfatter Krapina og Vindija i Jugoslavien; grottepladserne Kebara, Amud og Tabun i Israel; Shanidar i Irak; og den førnævnte Teshik Tash i Usbekistan.
De israelske lokaliteter har været af interesse i årtier, fordi de tilsyneladende er samtidige med nærliggende AMH-lokaliteter. Der har været mange spekulationer om arten af interaktioner mellem de to arter. En teori er, at da indlandsisen dækkede store dele af Europa, flyttede neandertalerne ned i Mellemøsten sammen med andre dyr. Det faktum, at AMH nåede Mellemøsten ved 120 kya, men aldrig kom ind i Europa før efter 40 kya, tyder for nogle på, at neandertalerne “holdt” Europa og forhindrede AMH i at trænge ind i Europa. Da neandertalerne bevægede sig ned i Mellemøsten, kan de have skubbet de herboende AMH ud af området. I de efterfølgende varmere perioder kan AMH have bevæget sig tilbage til området efter neandertalerne havde trukket sig tilbage til mere nordlige destinationer. Denne idé om byttepladser er nu blevet afløst af idéen om samtidighed og krydsning, i det mindste af nogle grupper på et eller flere tidspunkter.
Figur 35.2 illustrerer neandertalerens brede geografiske udbredelse. Det er sandsynligt, at populationerne under istidens fremrykning flyttede sig sydpå, således at populationerne i Vesteuropa var tættere på Middelhavet, og de østlige neandertaler kan have skubbet sig ned i Israel og andre varme områder sammen med andre dyr. De fossile optegnelser viser, at dyreflokke flyttede op og ned i breddegrader i overensstemmelse med klimatiske impulser, så det er meget sandsynligt, at homininpopulationer også gjorde det. De var kloge og kan have arvet tidligere kulturel viden, hvis de havde sprog og tanketeori, og om ikke andet havde de brug for at spise og ville have fulgt vildtet.
På det tidspunkt, hvor AMH flyttede ind i Vesteuropa ~35 kya, var neandertalerne begyndt at uddø. De bukkede sandsynligvis under for det stadig hårdere klima. De gennemgik også en evolutionær flaskehals på et tidspunkt og mistede noget af deres genetiske mangfoldighed, hvilket muligvis gjorde dem mere sårbare over for sygdomme. Ligesom i Mellemøsten har der været mange spekulationer om, hvad der skete, da AMH ankom til Vesteuropa. Selv om de sandsynligvis bar neandertalgener (medmindre disse vestlige AMH ikke efterlod nogen moderne efterkommere), har de måske ikke selv parret sig med neandertalerne, og de vestlige populationer ville helt sikkert have set noget anderledes ud end neandertalerne i Mellemøsten. De fleste østlige neandertaler var imidlertid forsvundet på det tidspunkt, hvor AMH passerede gennem deres tidligere østlige geografiske område på vej til Vesteuropa. Det er blevet foreslået, at AMH udkonkurrerede dem enten direkte, hvilket er kendt som konkurrencekonkurrence, eller indirekte, hvilket er kendt som scramble-konkurrence, eller måske endda dræbte dem, når de mødte dem. Konkurrence indebærer, at en gruppe forhindrer en anden gruppe i at få adgang til ressourcer, mens konkurrence indebærer, at en gruppe er bedre til at få adgang til ressourcerne end den anden gruppe. Jeg tænker altid på en bølle, der forsvarer et buffetbord mod andre, i modsætning til børn, der kæmper om at finde æg på en påskeægsjagt, hvor nogle er bedre end andre til at komme til og/eller finde æg. Det er også blevet bredt accepteret, at neandertalerne blev marginaliseret, da AMH trængte ind på deres territorium. Bortset fra en nyere dato fra den kroatiske lokalitet Vindija (28 kya) er de seneste datoer fra Den Iberiske Halvø, hvor man mener, at de trak sig tilbage og uddøde. Uanset hvad der skete mellem de to arter, da det ser ud til, at neandertalerne var på vej ud, er det sandsynligvis et uvedkommende spørgsmål. Det er ret passende, at efter alle de år, hvor man har troet, at mennesket spillede en rolle i neandertalerens undergang, ser det ud til, at de lavede kærlighed og ikke krig (i hvert fald så vidt vi ved)!
FYSISKE KARAKTERISTIK
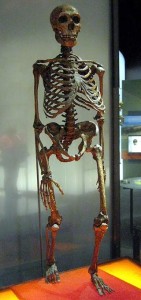
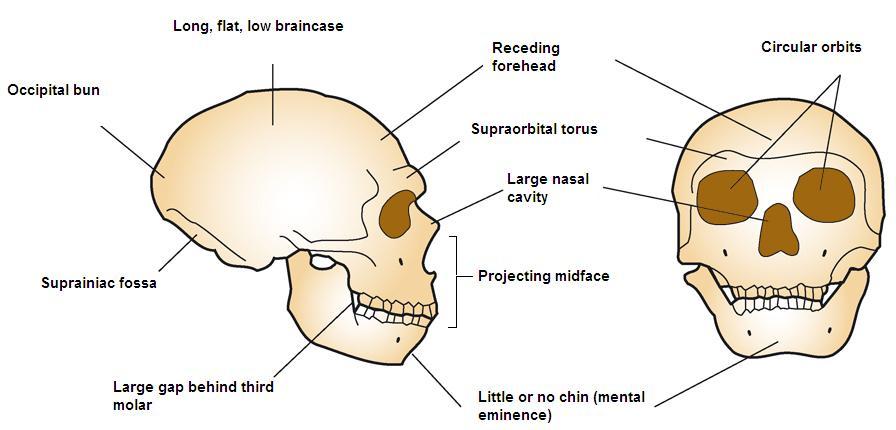
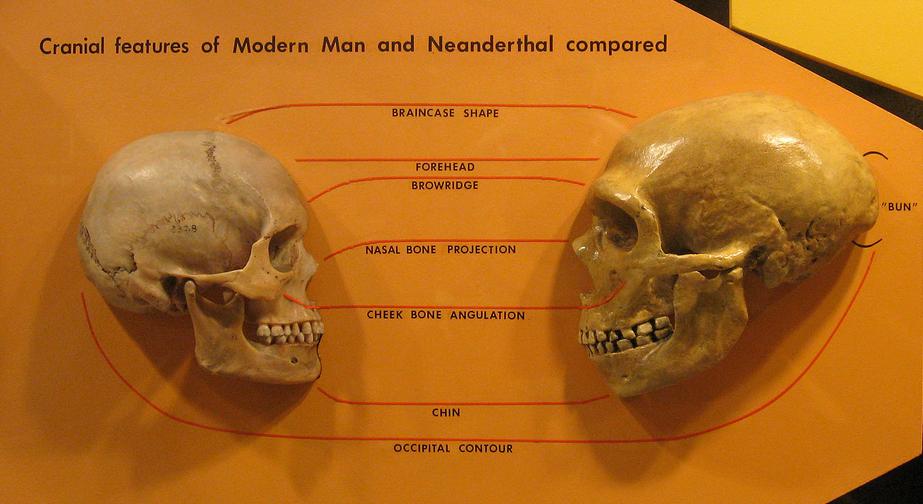
Se figur 35.4 for et komplet skeletbillede af en neandertaler. Som nævnt adskilte vestlige og østlige neandertaler sig morfologisk over tid. Denne kliniske variation, dvs. en gradueret ændring i fysiske karakteristika over det geografiske rum, menes at have været et resultat af tid, varierende tilpasning og eksponering for kronisk kolde istidsforhold og muligvis genstrøm med AMH i Mellemøsten. Populationer i Vesteuropa levede på højere breddegrader, og de klassiske neandertaler udviste kolde tilpasninger, der er i overensstemmelse med Bergmann’s og Allen’s regler. Bergmanns regel siger, at når man bevæger sig væk fra ækvator, øges massen i forhold til overfladearealet for at bevare varmen, da varmetabet er en funktion af overfladearealet. Allen’s regel vedrører længden af lemmer eller ekstremiteter, således at organismer i koldere miljøer har kortere vedhæng. I det ækvatoriale Afrika, hvor mennesker på lang sigt har tilpasset sig varme og tørre forhold, er kroppens morfologi således lang og gracile i modsætning til den korte, kraftige morfologi hos arktiske folkeslag. Ud over deres kraftige kroppe, korte vedhæng og tøndeformede brystkasse havde neandertalerne også ansigtstilpasninger til kulden. Ligesom H. heidelbergensis udviste neandertalerne midfacial prognathisme, store næser og oppustede ansigter på grund af forstørrede bihuler. På grund af deres fremadrettede overkæbe bevægede underkæben sig også fremad, hvilket efterlod et rum bag den tredje kindtand, et såkaldt retromolært rum. De indre næsefremspring var store, hvilket øgede det indre overfladeareal yderligere til opvarmning og befugtning af den indåndede luft. Desuden strakte unikke fremspring sig fra deres indre næseområde op i øjenhulerne. Yderligere kraniekarakteristika, der blev set hos både koldt tilpassede og østlige neandertaler, var store, jævnt rullede brynkamme over store, runde, vidtfordelte øjenhuler; “tilbagesvungne” zygomatikker; nogle unikke karakteristika ved det indre øre; og i baghovedregionen en baghovedbolle og suprainiac fossae (to små fordybninger placeret over inion, eller den ydre baghovedprotuberance; se Figur 35.5 for det generelle område). Mens deres kranier var længere og lavere end AMH’s kranier (se Figur 35.6), oversteg deres absolutte kraniekapacitet selv moderne menneskers kraniekapacitet. I overensstemmelse med Bergmanns regel er en større hjerne, selv om den er energimæssigt dyr i form af kalorier, mere konservativ ud fra et perspektiv med hensyn til varmeproduktion og -opbevaring. Mens neandertalernes hjerne var større, var AMH’s frontallapper og parietallapper (involveret i højere tankeprocesser) udvidet i forhold til neandertalerne. Dette kan have givet AMH en fordel i istidens Europa.
Postkranielt er neandertalerne blevet beskrevet som en krydsning mellem en maratonløber (med hensyn til deres udholdenhed) og en wrestler. De var bygget til at jage og dræbe byttedyr. Deres overkrop var stærkt muskuløs.
ENVIRONMENT AND WAY OF LIFE
Neandertalerne er traditionelt blevet portrætteret som havende udholdt barske klimatiske forhold. Der er imidlertid debat om, hvor stor en tolerance de havde over for forholdene i istidens Europa. Tattersall (2009) præsenterer en oversigt over forskning, der tyder på, at neandertalerne i de koldeste perioder levede i mere sydlige regioner og kun flyttede ind på højere breddegrader, når temperaturerne var varmere. Mens deres morfologi afspejler kronisk udsættelse for kulde, ligesom traditionelle arktiske folkeslag, havde de ikke den moderne teknologi, som disse moderne mennesker har, såsom bedre husly, skræddersyet tøj, våben og årtusinder af avancerede kulturelle traditioner. De var derfor nødt til at tilpasse sig biologisk til kulden. Men da forholdene forværredes, da det sidste istidsmaksimum nærmede sig, uddøde de sammen med andre dårligt tilpassede arter på de nordlige breddegrader.

Det har i mange år været kendt, at der var to forskellige typer neandertal-bopladser, og diskussionerne var ofte forudindtaget, som om nogle beboede førsteklasses ejendomme, mens andre ernærede sig på de åbne sletter. Hulepladser i Sydfrankrig blev ofte beskrevet som velvandede floddale med rigeligt med huler til skjul, vildt, vand, stenressourcer til redskaber osv. og man mente, at de havde været beboet i årtusinder. Steder i det åbne land var tilsyneladende beboet af mere nomadiske grupper, der levede i det fri i fritstående strukturer og fulgte efter flokdyr. Man mener nu, at grottepladser og friluftspladser repræsenterer det sæsonbestemte skift i subsistensstrategier, der blev praktiseret af de samme folkeslag.
Så mens neandertalerpopulationer kan have været i stand til at leve året rundt i mere tempererede områder, var resten sandsynligvis semi-nomadiske som deres forfædre. Under den kolde vinter i Vesteuropa søgte de sandsynligvis ly i sydlige huler og om sommeren vovede de sig nordpå i jagten på vandrende flokke. Ud over huler brugte de også klippeskjul, hvor de byggede ud fra en klippevæg eller et overhæng. De gjorde det samme inde i grotterne, idet de byggede et skjul i et skjul. Stolpeforme dannes, når en træstolpe, der er sat ned i jorden, forfalder, så man kan se en mørkere cirkel af humus. Mønsteret af stolpeforme kan bruges til at rekonstruere form og størrelse af huse og vægge i arkæologiske optegnelser. På baggrund af stolpeforme ved man, at neandertalerne har dækket huleindgange, sandsynligvis i koldere perioder. Rester af konstruerede strukturer viser, at de brugte knogler, stolper og sten og sandsynligvis dækkede dem med skind og isolerede dem med græs. På Moldova-området i Ukraine blev der udgravet en 26 x 16´ oval ring af mammutknogler. Knoglerne var sandsynligvis dækket af skind og dannede en hytte, der indeholdt mange ildsteder.
I løbet af Pleistocæn beskrives det, at de europæiske vintre var lange og kolde, og at somrene var korte og kølige. På grund af sæsonafhængigheden ville plantefødevarer primært have været tilgængelige i de varmere måneder. De europæiske neandertaler spiste en stor andel af kød, hvor rensdyr og mammut udgjorde størstedelen af kosten, baseret på henholdsvis faunasamlinger og isotopanalyser. Kostsammensætningen varierede dog fra region til region. Heste, kvæg og geder levede på sletterne, mens bjergfår og stenbukke dominerede i de højere beliggende områder. På stedet Shanidar i Irak omfattede faunaresterne knogler af geder, får, kvæg, svin, skildpadder, bjørne, rådyr, ræve, mårdyr og gerbiler. På samme sted er der beviser for planteforbrug og madlavning. Henry (2011) fandt phytolitter og stivelseskorn i kalkaflejringer (calculus) på neandertaltænder. Noget af stivelsen fra græsser viste skader, der er karakteristiske for kogning. Selv om vi ved, at neandertalerne brugte ild, som det fremgår af ildsteder på deres lokaliteter, og at de sandsynligvis spiste planter, når de var tilgængelige, er det værdifuldt endelig at have understøttende beviser. Da Shanidar ligger syd for det meste af Europa og dermed er mere tempereret, er det sandsynligt, at neandertalerne havde større adgang til sådanne ressourcer.

Mens debatten i nogen tid har raset om, hvorvidt neandertalerne praktiserede kannibalisme, giver fossilt materiale, især fra den franske lokalitet Moula-Guercy, overbevisende beviser for, at i det mindste nogle grupper spiste deres egne. Neandertalsknogler på stedet viser de samme tegn på forarbejdning som dyreknogler. Knoglerne er blevet skilt ad og hamret op for at finde marv, og de viser snitmærker fra fjernelse af muskler.
Det er interessant, hvor afskyeligt kannibalisme er for os. Vi identificerer os med neandertalerne og kan føle os skuffede over, at de praktiserede kannibalisme. Selv om det er svært at sige, hvorfor de spiste hinanden, er der tegn på koststress i form af emaljehypoplasi på nogle steder, f.eks. i Krapina i Kroatien. Således led nogle grupper af periodisk fødevaremangel, som resulterede i mangelfuld emaljeaflejring hos børn i udvikling. Hvis folk sulter, og der er et lig til rådighed, viser historiske beretninger, at de vil spise det. Det burde derfor ikke være overraskende, hvis neandertalerne spiste de døde i stedet for at dræbe dem for at spise dem. Der er ingen beviser for, at de praktiserede kannibalisme i alle tider og alle steder, og det kunne således have været en reaktion på ekstreme forhold.
For beviserne fra Moula Guercy var der stor modvilje, hver gang nogen foreslog kannibalisme som forklaring på skader på levn på bestemte steder. Jeg tror, at det, der stimulerer mange af os til at føle affinitet med neandertalerne, er, at de med vilje begravede deres døde. De virker så menneskelige. Nu hvor vi ved, at nogle af vores forfædre krydsede sig med dem, føler vi måske en endnu større forbindelse til dem og bliver nødt til at acceptere det gode sammen med mulige overlevelseshandlinger.

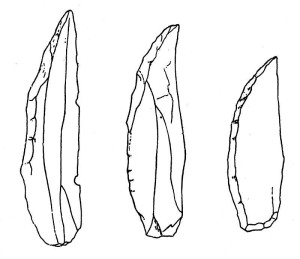
Neandertal-kulturen falder inden for den periode, der betegnes som den mellemste stenalder, dvs. den midterste del af den gamle stenalder. Den neandertale redskabstradition betegnes Mousterian Industry (se figur 35.9 og 35.10 for eksempler på Mousterian redskaber), efter Le Moustier-området i Frankrig (se figur 35.8). Selv om der ikke er nogen kendte neandertal-rester fra Nordafrika, er det interessant, at deres redskaber er blevet fundet der (se figur 35.11). Mousterian-metoden var en forbedring af Levallois-teknikken, der gav større kontrol over de resulterende flager. Flagerne blev derefter modificeret til en række forskellige redskaber, såsom skrabere og spidser, til forskellige funktioner. Nogle af værktøjerne var dentikulerede, hvilket betyder, at de var savtakkede. Ligesom H. heidelbergensis fremstillede de sammensatte redskaber ved at sætte stenredskaber på håndtag og skafter.

En senere værktøjstradition (35 kya) fra St. Cesaire-lokaliteten i Frankrig kategoriseres som en industri fra den øvre palæolitiske stenalder, da værktøjerne udviser karakteristika for AMH-industrier. Den kaldes Chatelperronian-traditionen (se figur 35.12) og kan være et bevis på direkte eller indirekte kontakt mellem neandertalerne og AMH, hvilket betyder, at de fik teknologien via kontakt med AMH eller fandt et eller flere redskaber og brugte deres egne metoder til at efterligne dem. Nogle forskere hævder det sidstnævnte versus indlæring af AMH’s produktionsmåder. Adskillige genstande fra neandertalstedet Arcy-sur-Cure, Frankrig, er blevet fortolket som smykker, en anden kulturel præstation, der udelukkende tilskrives AMH.
Mens H. naledi og H. heidelbergensis deponerede deres døde i dybe huler, var neandertalerne den første art, der vides at begrave deres døde i individuelle grave. Kroppene blev ofte fundet i en bøjet stilling. Der er meget få tegn på ritualer i forbindelse med neandertalbegravelser. Det ser ud til, at de gravede et hul, foldede liget ned i hullet, deraf den bøjede stilling, og muligvis smed nogle andre ting i hullet sammen med det. Genstande fortolkes ofte som havende en vis betydning, men de er normalt begrænset til dyreknogler og knuste redskaber. På stedet Teshik Tash i Usbekistan blev en niårig dreng imidlertid begravet med fem sæt vilde gedehorn, som muligvis prydede hans krop. Mens nogle har foreslået, at han var en AMH, synes det, hvis han var neandertaler, at have været en ritualiseret begravelse.
Shanidar-lokaliteten (se figur 35.13) har altid været den mest romantiske fra mit perspektiv. Det er en grotteplads, der oplevede periodiske huleindbrud, og som har givet resterne af flere interessante individer, hvoraf nogle blev begravet med vilje. Shanidar 1 var en voksen mand. Selv om han i sidste ende blev offer for et huleindbrud, overlevede han en eller flere tidligere traumatiske hændelser i sit liv. Det menes, at han var delvis blind på grund af en hovedskade, der involverede et af hans øjne. Han manglede enden af en af sine underarme og dermed også hånden. Han led en skade på benet, der resulterede i en permanent halthed, og nogle af hans tænder var helt nedslidte. Det interessante spørgsmål er, hvordan han overlevede? Det ofte citerede svar er, at hans gruppekammerater hjalp ham i livet. Han er således udråbt som endnu et tilfælde af førmenneskelig altruisme eller i det mindste slægtsvalg, hvis plejen blev ydet af hans slægtninge.
Shanidar 3 er måske det tidligste bevis for mord. Han var også en voksen mand og blev muligvis stukket ned, hvilket et snitsår på et af hans ribben vidner om. Selv om knoglen viste tegn på heling, vides det ikke, om han døde af såret og blev begravet med vilje eller om han døde i et huleindbrud. En anden interessant begravelse er Shanidar 4. Han var også en voksen mand, der blev begravet med vilje, og der blev fundet pollen fra otte arter af vilde blomster i graven. Selv om det er meget kontroversielt og muligvis kan tilskrives en gnaverart, der gemmer frø, vil mange gerne tro, at neandertalerne ikke blot begravede deres døde, men også lagde blomster på deres rester.
Mens jeg skrev det foregående afsnit, slog det mig, at hanner måske er blevet begravet forskelligt i forhold til hunnerne. Selv om jeg ikke kender svaret på det spørgsmål, er det interessant og stof til eftertanke.

Debatten om, hvorvidt neandertalerne kunne tale, har raset i årtier. I mange år mente eksperterne, at deres strubehoved lå for højt oppe i halsen til, at de kunne tale. Vores strubehoved falder i løbet af udviklingen. Tidligt er det placeret højt oppe i halsen for at muliggøre samtidig at drikke og trække vejret. Spædbørn kan ikke tale, før strubehovedet falder ned, og de begynder derefter at pludre. Selv om de således ikke er mekanisk i stand til at tale tidligt, er de kognitivt i stand til at lære sprog. Det er interessant, at nogle forældre nu lærer deres spædbørn tegnsprog, så de kan kommunikere tidligere.
Mens nogle forskere stadig tvivler på neandertalerne evne til at tale, har mange accepteret, at de sandsynligvis havde talesprog, men at de ikke ville have været i stand til at producere hele den række af lyde, der karakteriserer vores egen tale. Opdagelsen af et neandertalsk tungeben på Kebara-området i Israel fik mange til at acceptere deres evne til at tale, da dets morfologi lignede vores egen. Hyoidknoglen er et vigtigt fastgørelsessted for ledbånd og brusk i strubehovedet og for nogle af tungens extrinsiske muskler (f.eks. geniohyoid, hyoglossus). Det mest afslørende bevis til støtte for neandertalernes tale er, ud over alle mine tidligere argumenter, tilstedeværelsen af FOX P2-genet i deres genom. Vi besidder også genet, og det spiller en vigtig rolle i tilegnelsen af sprog.