Introduktion
Gram-positive bakterier af Bacillus- og Clostridium-slægterne kan differentiere sig for at danne en endospore (spore), en metabolisk hvilende celle, der produceres som reaktion på forhold, der ikke tillader cellevækst (McKenney et al., 2012). Når sporen først er frigivet i miljøet, overlever den i sin hvilende tilstand i ekstremt lange perioder og er modstandsdygtig over for en lang række stressfaktorer som f.eks. høje temperaturer, dehydrering, mangel på næringsstoffer og tilstedeværelse af giftige kemikalier (McKenney et al., 2012). Den hvilende spore er imidlertid i stand til løbende at overvåge miljøet og reagere på tilstedeværelsen af vand og næringsstoffer ved at spire og frembringe en vegetativ celle, der er i stand til at vokse og sporulere (McKenney et al., 2012). Resistens over for ikke-fysiologiske forhold skyldes til dels sporernes overfladestrukturer. I Bacillus subtilis, modelsystemet for sporedannere, er sporeoverfladen organiseret i en flerlaget kappe og i en skorpe, et nyligt opdaget lag, der omgiver sporekappen (McKenney et al., 2012). B. subtilis-sporer er negativt ladede (Huang et al., 2010; Pesce et al., 2014) og har en relativ hydrofobicitet, som til dels skyldes glykosylering af nogle sporeoverfladeproteiner (Cangiano et al., 2014; Rusciano et al., 2014). Hos flere Bacillus- og Clostridium-arter, herunder B. cereus, B. anthracis, B. megaterium og C. difficile, er den yderste sporestruktur exosporium, et morfologisk særskilt lag bestående af proteiner og glykoproteiner, der omgiver kappen (Díaz-González et al., 2015; Manetsberger et al, 2015b; Stewart, 2015).
Bakteriesporen er blevet foreslået som en platform til at vise heterologe proteiner, med potentielle anvendelser, der spænder fra udvikling af vacciner til slimhinder til genanvendelige biokatalysatorer, diagnostiske værktøjer og bioremedieringsanordninger (Knecht et al., 2011; Isticato og Ricca, 2014; Ricca et al., 2014). Sporens bemærkelsesværdige og veldokumenterede resistens (McKenney et al., 2012), flere sporedannende arters modtagelighed for genetisk manipulation (Harwood og Cutting, 1990) og flere arters sikkerhed (Cutting, 2011; Baccigalupi et al., 2015) understøtter brugen af sporerne som et display- og leveringssystem. Der er blevet udviklet to strategier til at vise heterologe proteiner på sporens overflade. En rekombinant strategi, der er baseret på konstruktion af genfusioner mellem DNA, der koder for et udvalgt sporeoverfladeprotein, og DNA, der koder for det protein, der skal vises, er blevet anvendt i årenes løb til at vise en række heterologe proteiner (Isticato og Ricca, 2014). En ikke-rekombinant tilgang, der er baseret på spontan adsorption mellem rensede sporer og rensede proteiner, er også blevet anvendt til at vise forskellige enzymer og antigener (Ricca et al., 2014). De molekylære detaljer, der styrer sporeadsorptionen, er ikke blevet fuldt ud belyst. Det er kendt, at adsorptionen er mere effektiv, når pH-værdien af bindingsbufferen er sur (pH 4) (Huang et al., 2010; Sirec et al., 2012), og at en kombination af elektrostatiske og hydrofobiske interaktioner sandsynligvis er involveret i interaktionen (Huang et al., 2010; Sirec et al., 2012). Det er også kendt, at muterede sporer med alvorligt ændrede sporeoverflader interagerer mere effektivt end isogene vildtypesporer med modelproteiner (Sirec et al., 2012; Donadio et al, 2016).
Her brugte vi et fluorescerende protein, den monomere form af det røde fluorescerende protein (mRFP) fra korallen Discosoma sp. (Campbell et al., 2002), til at evaluere, om sporer af B. megaterium er i stand til at interagere med og adsorbere et heterologt modelprotein. B. megaterium omfatter en række morfologisk forskellige stammer, der deler de usædvanligt store dimensioner af både celler (længde på op til 4 μm og en diameter på 1,5 μm) og sporer (længde på op til 3 μm og en diameter på 1 μm) (Di Luccia et al., 2016). Sporer af nogle stammer af B. megaterium er omgivet af et exosporium, og da hidtil kun sporer af arter uden exosporium er blevet betragtet som adsorptionsplatforme, foreligger der ingen data om exosporiets indvirkning på interaktionen med heterologe proteiner.
Den QM B1551-stamme er den bedst karakteriserede stamme af B. megaterium. Denne stamme bærer omkring 11% af sit genom på syv indfødte plasmider (Rosso og Vary, 2005; Vary et al., 2007; Eppinger et al., 2011), hvoraf to – pBM500 og pBM600 – er blevet identificeret som bærere af gener, der er essentielle for dannelsen af denne stammes karakteristiske “valnøddeformede” exosporium (Manetsberger et al., 2015a). Proteinsammensætningen af QM B1551 exosporium er endnu dårligt karakteriseret, idet kun nogle få gener, der koder for ortologer af anerkendte exosporieproteiner i sporer af andre arter, er identificeret ved genomiske analyser. Disse omfatter gener, der koder for BclA nap-proteiner, som danner en lokaliseret nap i B. megaterium QM B1551-sporer, samt en ortolog af BxpB-proteinet, der danner det basale lag af exosporiet i sporer fra B. cereus-familien. Sidstnævnte synes at udfylde en anden rolle i B. megaterium QM B1551-sporer, da en nulmutantstamme bevarede et tilsyneladende normalt exosporium (Manetsberger et al., 2015a).
I denne artikel præsenterer vi data, der viser, at sporer af B. megaterium QM B1551 effektivt kan adsorbere mRFP, og giver beviser for, at proteinmolekyler er i stand til at krydse den permeabilitetsbarriere, som exosporiumet udgør, for at lokalisere sig i intercoat-rummet.
Materialer og metoder
Bakteriestammer, sporer og RFP-produktion
De B. megaterium-stammer, der er anvendt i denne undersøgelse, er QM B1551 og dens plasmidløse derivat PV361 (Rosso og Vary, 2005). Den B. subtilis-stamme, der blev anvendt i denne undersøgelse, var PY79 (Youngman et al., 1984). Sporulation af alle Bacillus-stammer blev induceret ved hjælp af udmattelsesmetoden (Cutting og Vander Horn, 1990). Efter 30 timers vækst i Difco Sporulation (DS)-medium ved 37°C med kraftig rystning blev sporer indsamlet og renset som beskrevet af Nicholson og Setlow (1990) ved hjælp af inkubation natten over i H2O ved 4°C for at lyse resterende sporangialceller. Antallet af rensede sporer blev målt ved direkte optælling med et Bürker-kammer under et optisk mikroskop (Olympus BH-2 med 40× linse).
Til produktion af mRFP blev celler af Escherichia coli stamme RH161 (Donadio et al, 2016), der bærer ekspressionsvektoren pRSET-A, der bærer en in-frame-fusion af 5′-enden af den rfp-kodende region til seks histidin-kodoner under transkriptionskontrol af en T7-promotor, blev dyrket i 18 timer ved 37 °C i 100 ml autoinduktionsmedium for at udtrykke det heterologe protein (Isticato et al., 2010). Det His6-mærkede RFP-protein blev oprenset under native betingelser ved hjælp af en His-Trap-kolonne som anbefalet af producenten (GE Healthcare Life Science). Oprenset protein blev afsaltet ved hjælp af en PD10-kolonne (GE Healthcare Life Science) for at fjerne høje NaCl- og imidazolkoncentrationer.
Adsorptionsreaktion
Medmindre andet er angivet, blev 5 μg oprenset rekombinant mRFP tilsat til en suspension af sporer (5 × 108) i 50 mM natriumcitrat pH 4,0 ved 25 °C i et slutvolumen på 200 μl. Efter 1 times inkubation blev bindingsblandingen centrifugeret (10 min. ved 13.000 g) for at fraktionere mRFP-bundne sporer i pellen fra fri mRFP i supernatanten.
Western- og Dot-Blot-analyse
Sporepiller fra adsorptionsreaktioner blev resuspenderet i 40 μl sporecoat-ekstraktionsbuffer (Nicholson og Setlow, 1990; Giglio et al, 2011), inkuberet ved 68°C i 1 h for at solubilisere sporehindeproteiner og indlæst på en 12% SDS-PAGE-gel. Proteinerne blev derefter elektrotransporteret til nitrocellulosefiltre (Amersham Pharmacia Biotech) og brugt til Western Blot-analyse som tidligere rapporteret (Isticato et al., 2008) ved hjælp af monoklonalt mRFP-genkendeligt anti-His-antistof (Sigma). En kvantitativ bestemmelse af mængden af mRFP blev opnået ved dot blot-forsøg ved at sammenligne serielle fortyndinger af renset mRFP og bindingsassay-supernatant. Filtrene blev derefter visualiseret ved hjælp af ECL-prime-metoden (Amersham Pharmacia Biotech) og underkastet densitometrisk analyse ved hjælp af Quantity One 1-D Analysis Software (Bio-Rad). Dot blot- og relative densitometriske analyser blev udført tre gange for at verificere resultaternes signifikans.
Fluorescens- og immunofluorescensmikroskopi
Post-adsorptionssporer blev resuspenderet i 50 μl 1x PBS pH 4,0, og 5 μl af suspensionen blev anbragt på mikroskopglas og dækket med et dækglas, der forinden var behandlet i 30 s med poly-L-lysin (Sigma). Immunofluorescens blev udført som beskrevet af Isticato et al. (2013), med følgende ændringer: 2,0 × 106 RFP-adsorberede sporer af QM B1551 og PV361 B. megaterium-stammer blev forbehandlet med 1% bovin serumalbumin (BSA) – 1x PBS pH 4,0 i 30 min forud for 2 h-inkubering ved 4°C med anti-polyHistidin-antistoffer (mus; Sigma) fortyndet 1:20 i 1x PBS pH 4,0-1% BSA. Som en kontrol af denne tekniks specificitet blev ikke-adsorberede sporer direkte behandlet med anti-His-antistoffer. Efter tre vaske blev prøverne inkuberet med et 64-foldigt fortyndet sekundært anti-mus-antistofkonjugeret med fluoresceinisothiocyanat, FITC (Santa Cruz Biotechnology, Inc.) og vasket fire gange med 1x PBS pH 4,0. Vaskede prøver blev resuspenderet i 20 μl 1x PBS pH 4,0, og 10 μl blev analyseret. Alle prøver blev observeret med et Olympus BX51 fluorescensmikroskop udstyret med et 100× UPlan F1 olieobjektiv; der blev anvendt U-MNG- eller U-MWIBBP-kubefiltre til at detektere henholdsvis den røde fluorescensemission af mRFP eller den grønne emission af FITC-konjugerede antistoffer. Eksponeringstiden var mellem 200 og 3000 ms. Billeder blev optaget ved hjælp af et Olympus DP70 digitalkamera og behandlet med Image Analysis Software (Olympus) til mindre justeringer af lysstyrke, kontrast og farvebalance (McCloy et al., 2014). ImageJ (v1.48, NIH) blev brugt til at tegne en omrids omkring 80 sporer for hver stamme, og minimum, maksimum og gennemsnitlige fluorescensværdier pr. pixel blev registreret for hver spore. Værdierne for fluorescensintensitet blev efterfølgende vist som box-plots med 5-95 % konfidensintervaller (McCloy et al., 2014).
Statistisk analyse
Resultaterne fra dot blot- og fluorescensmikroskopianalyse er gennemsnit fra tre uafhængige eksperimenter. Statistisk signifikans blev bestemt ved Student t-test, og signifikansniveauet blev sat til P < 0,05.
Resultater
mRFP af Discosoma sp. er Adsorberet af B. megaterium Sporer
For at verificere, om sporer af B. megaterium QM B1551 var i stand til at adsorbere mRFP, blev 5 μg af det oprensede protein (Materialer og metoder) inkuberet med 5,0 × 108 oprensede sporer. Adsorptionsreaktionen blev udført i 50 mM natriumcitrat ved pH 4,0, som tidligere beskrevet (Sirec et al., 2012). Efter reaktionen blev sporer vasket grundigt med 1x PBS pH 4,0, sporeoverfladeproteiner blev ekstraheret som beskrevet i Materialer og metoder og analyseret ved western blotting med anti-polyHistidine-Peroxidase-antistof (Sigma), som genkender den histidine-tagged N-terminus af mRFP. Som vist i figur 1A blev mRFP ekstraheret fra sporer, hvilket indikerer, at det blev absorberet under reaktionen og derefter frigivet ved ekstraktionsbehandlingen. For at vurdere adsorptionens effektivitet analyserede vi mængden af mRFP, der blev efterladt ubundet, dvs. efter adsorberede sporer blev de opsamlet ved centrifugering, og supernatanten blev fortyndet serielt og analyseret ved dot blotting (figur 1B). En densitometrisk analyse af dotblot (Supplerende tabel 1) viste, at når 5 μg mRFP blev anvendt i adsorptionsreaktionen, blev mindre end 1 % efterladt ubundet, hvilket indikerer, at omkring 99 % af det heterologe protein blev adsorberet til B. megaterium-sporer.

FIGUR 1. Bacillus megaterium QM B1551-sporer adsorberer mRFP. 5 × 108 sporer blev inkuberet med 5 μg oprenset mRFP, og derefter blev prøverne centrifugeret. (A) Sporeoverfladeproteiner blev ekstraheret fra pelletfraktionen ved SDS-DTT-behandling, fraktioneret på SDS-PAGE og analyseret ved Western Blot. Renset mRFP (20 μg) blev anvendt som markør. (B) Supernatanten, der indeholdt den ubundne mRFP, blev fortyndet serielt og analyseret ved dot blot (QM B1551). Serielle fortyndinger af renset mRFP (mRFP) blev anvendt som standard. (C) Sporer adsorberet med mRFP blev vasket to gange (W1 og W2) med PBS-buffer ved pH 3,0 eller pH 7,0 eller med en 1M NaCl, 0,1% Triton X-100-opløsning. Der blev anvendt serielle fortyndinger af renset mRFP og ubundet mRFP (UN-mRFP) som standarder. Immunreaktioner i alle paneler blev udført med mRFP-genkendeligt anti-His-antistof konjugeret med peberrodsperoxidase.
For at analysere, om adsorberede mRFP-molekyler var tæt bundet til sporeoverfladen, blev sporer efter adsorptionsreaktionen vasket to gange med PBS-buffer ved pH 3,0 eller pH 7,0 eller med en 1M NaCl, 0,1% Triton X-100-opløsning som tidligere beskrevet (Donadio et al., 2016). Som vist i Figur 1C, og understøttet af den densitometriske analyse af dot blot (Supplerende tabel 2), forårsagede vaskningerne ved pH 3,0 ikke nogen frigivelse af de adsorberede mRFP, mens vaskningerne ved pH 7,0 eller med 1M NaCl, 0,1% Triton X-100 forårsagede en minimal, mindre end 1%, frigivelse af mRFP-molekyler. Resultaterne i figur 1 tyder derfor på, at mRFP blev adsorberet effektivt og var tæt bundet til B. megaterium-sporer. For at vurdere, om spor-adsorberede mRFP-molekyler bevarede deres fluorescensegenskaber, udførte vi en fluorescensmikroskopianalyse. Som vist i figur 2 var sporer efter adsorptionsreaktionen forbundet med et stærkt fluorescenssignal, der var synligt omkring hele sporeoverfladen.
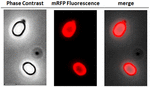
FIGUR 2. Fluorescensmikroskopianalyse af B. megaterium QM B1551-mRFP-sporer. QM B1551-sporer, der var inkuberet med mRFP (5 μg) og efterfølgende vasket, blev analyseret ved fluorescensmikroskopi. Det samme mikroskopifelt blev observeret med fasekontrast- og fluorescensmikroskopi. Skala bar 1 μm. Sammenlægningspanelet er rapporteret. Eksponeringstiden var 200 ms.
Exosporium har en afgørende rolle i mRFP-adsorption
Stamme QM B1551 af B. megaterium indeholder syv indfødte plasmider (Rosso og Vary, 2005; Eppinger et al., 2011), og plasmidkodede gener er afgørende for exosporiumdannelse (Manetsberger et al., 2015a). PV361 er en QM B1551-hærdet stamme, der mangler alle syv plasmider, og som følge heraf mangler den helt exosporium (Manetsberger et al., 2015a). Vi brugte sporer af stamme PV361 til at analysere exosporiets rolle i mRFP-adsorptionen. Parallelt hermed brugte vi også sporer af B. subtilis PY79, som i en tidligere undersøgelse er blevet vist at adsorbere mRFP (Donadio et al., 2016). For at sammenligne adsorptionseffektiviteten af sporer af B. subtilis PY79- og B. megaterium QM B1551- og PV361-stammerne adsorberede vi 5 μg oprenset mRFP med 5,0 × 108 sporer af hver af de tre stammer. Efter adsorptionsreaktionerne blev sporer opsamlet ved centrifugering, proteiner blev ekstraheret ved SDS-DTT-behandling og analyseret ved western blotting med mRFP-genkendeligt anti-His-antistof. Som det fremgår af figur 3A, blev mRFP tilsyneladende ekstraheret i større mængder fra sporer af QM B1551 end fra sporer af de to andre stammer. I et forsøg på at kvantificere disse tilsyneladende forskelle blev ubundet mRFP fra adsorptionsreaktionerne fortyndet serielt og analyseret ved dot blotting (figur 3B). En densitometrisk analyse af dotblot af figur 3B (Supplerende tabel 3) viste, at PY79- og PV361-sporer adsorberede omkring 90 % af den samlede mRFP, mens QM B1551-sporer adsorberede næsten al (over 99 %) renset mRFP.
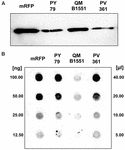
FIGUR 3. Monomerisk rødt fluorescerende protein adsorption til sporer af B. megaterium QM B1551 og PV361 og B. subtilis PY79. 5 × 108 sporer af hver stamme blev inkuberet med 5 μg oprenset mRFP, hvorefter prøverne blev centrifugeret. Sporer i pelletfraktionerne blev brugt til at ekstrahere overfladeproteiner, som efterfølgende blev analyseret ved western blot (A), mens supernatanterne blev fortyndet serielt og analyseret ved dot blot (B). Serielle fortyndinger af renset mRFP blev anvendt som standarder. Immunreaktioner i begge paneler blev udført med anti-His-antistof konjugeret med peberrodsperoxidase.
Adsorptionseffektiviteten af sporer af de tre stammer blev også analyseret ved fluorescensmikroskopi (Figur 4A og Supplerende figur S1). Mikroskopibilleder blev analyseret af ImageJ-software (v1.48, NIH) for at udføre kvantitativ fluorescensbilledeanalyse, og sporefluorescens blev beregnet som beskrevet i Materialer og metoder. Analysen af 80 sporer af hver stamme viste en gennemsnitlig fluorescensværdi pr. pixel på 52.43 (± 5.94), 20.81 (± 2.71) og 32.33 (± 2.97; arbitrære enheder) for henholdsvis QM B1551, PV361 og PY79 (figur 4B), hvilket giver yderligere bevis for, at QM B1551-sporer adsorberede mere mRFP end sporer fra de to andre stammer.
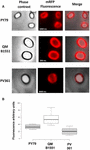
FIGUR 4. Effektiv adsorption af mRFP til sporer af B. megaterium QM B1551 og PV361 samt B. subtilis PY79. (A) Fluorescensmikroskopibilleder af PY79-, QM B1551- og PV361-sporer efter mRFP-adsorption og vask. Eksponeringstiden er angivet. Faskontrast og røde fluorescensoverlejringer er vist (flettepanel). Skalaen er 1 μm. (B) Boxplots med angivelse af fluorescensintensiteten af 80 forskellige sporer af hver stamme. Grænserne i hver boks repræsenterer den første og tredje kvartil (25 og 75 %), og værdierne uden for boksene repræsenterer maksimum- og minimumsværdierne. Den linje, der deler boksen, angiver medianværdien for hver stamme. P-værdien er mindre end 0,0001.
Baseret på resultaterne i figur 3 og 4 konkluderer vi, at det exosporium, der er til stede i QM B1551 og mangler i PV361, har en relevant rolle i adsorptionen af mRFP.
Dertil kommer, at resultaterne i figur 4 viste, at B. subtilis PY79-sporer er mere effektive end B. megaterium PV361-sporer til at adsorbere mRFP, mens dot blotting rapporteret i figur 3B viste lignende adsorptionseffektiviteter for de to stammer. Vi mener, at denne uoverensstemmelse skyldes en stærk reduktion af fluorescensen, når mRFP er bundet til PV361, men ikke til PY79- eller QM B1551-sporer (se nedenfor).
Kvantitativ vurdering af mRFP-adsorption til QM B1551-sporer
Dot blot-eksperimenter (figur 3B) viste, at når 5 μg oprenset mRFP blev anvendt i adsorptionsreaktioner med 5,0 × 108 sporer af QM B1551-stammen, blev næsten alt heterologt protein bundet til sporen. For at definere den maksimale mængde mRFP, der kan adsorberes til QM B1551-sporer, gentog vi reaktionerne med stigende koncentrationer af renset mRFP, dvs. at 5,0 × 108 QM B1551-sporer blev reageret med 5, 10, 20, 40, 80 og 160 μg renset mRFP. Efter reaktionerne blev sporer opsamlet ved centrifugering, og supernatanterne, der indeholdt ubundet mRFP, blev fortyndet serielt og analyseret ved dot blotting (figur 5A). Figur 5B viser resultaterne af en densitometrisk analyse af dotblot (??), som viser, at når 5-80 μg mRFP blev reageret med 5 × 108 sporer, var procentdelen af protein bundet til sporer over 90 %. Der blev observeret et fald i den bundne mRFP-andel, når 160 μg oprenset protein blev anvendt i reaktionen. Men selv når 160 μg renset mRFP blev anvendt, blev over 60 % af proteinet absorberet, hvilket indikerer, at 5,0 × 108 sporer af QM B1551 kan adsorbere ca. 100 μg mRFP.
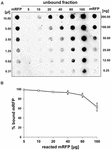
FIGUR 5. Kvantitativ vurdering af mRFP-adsorption til B. megaterium QM B1551-sporer. 5 × 108 sporer blev inkuberet med 5, 10, 20, 40, 80 og 160 μg renset mRFP. Reaktionsblandingerne blev efterfølgende centrifugeret, og supernatanterne blev fortyndet serielt og analyseret ved dot blot (A). Der blev anvendt serielle fortyndinger af oprenset mRFP som standarder. Immunreaktionerne i begge paneler blev udført med anti-His-antistof konjugeret med peberrodsperoxidase. (B) Procentdel af mRFP adsorberet til sporer efter reaktion med definerede mængder endogen mRFP. Fejlstregs viser standardfejlene af gennemsnittet fra tre eksperimenter og P-værdien aldrig over 0,0025.
mRFP lokaliseres til intercoat-rummet i B. megaterium QM B1551-sporer
En immunfluorescensmikroskopitilgang blev anvendt til at vurdere, om adsorberede mRFP-molekyler blev udsat på overfladen af B. megaterium QM B1551-sporer. QM B1551-sporer adsorberet med forskellige mængder mRFP blev reageret med monoklonalt anti-His-antistof, der genkender det rekombinante mRFP, og derefter med fluorescerende sekundært anti-mus-antistof (Santa Cruz Biotechnology, Inc.) og observeret under fluorescensmikroskopet (figur 6). Med den laveste mængde mRFP, der blev anvendt i dette forsøg (2 μg), blev mRFP-fluorescenssignalet (rødt) observeret hele vejen rundt om sporerne, mens immunofluorescenssignalet (grønt) var svagt og hovedsagelig koncentreret ved sporepolerne, hvilket tyder på, at mRFP kun var eksponeret på sporeoverfladen på disse steder. Antallet af grønne pletter steg med den mængde mRFP, der blev anvendt i reaktionen (5 og 10 μg), og med den største mængde mRFP, der blev anvendt i reaktionen (20 μg), blev der observeret en næsten fuldstændig grøn ring omkring sporerne. På grundlag af resultaterne i figur 6 antog vi, at mRFP-molekyler infiltrerer gennem exosporium og lokaliseres i mellemrummet mellem den ydre kappe og exosporium, dvs. at når der anvendes en lav mængde mRFP, er næsten alle proteinmolekyler internt i exosporium og er kun tilgængelige for antistoffet på få steder. Stigende mængder mRFP i adsorptionsreaktioner resulterer i, at mellemrummene mellem pelsene “fyldes op”, indtil der i sidste ende er flere mRFP-molekyler til rådighed for antistoffet på sporeoverfladen. Denne hypotese indebærer, at hvis der mangler exosporium, bør al mRFP være tilgængelig for antistoffet. For at teste dette sammenlignede vi ved immunofluorescensmikroskopi lige mange sporer af QM B1551 (med exosporium) og af PV361 (uden exosporium), som var inkuberet med den samme mængde mRFP (5 μg). Når exosporium var til stede (QM B1551), var mRFP kun delvist tilgængeligt for antistoffet, og der blev observeret grønne pletter (figur 7 og supplerende figur S2). Når exosporiet ikke var til stede (PV361), var adsorberet mRFP tilgængeligt for antistoffet hele vejen rundt om sporen, og der blev dannet en komplet grøn ring, hvilket understøtter hypotesen om, at mRFP er internt i exosporiet i QM B1551-sporer.
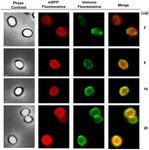
FIGUR 6. Immunofluorescensanalyse af mRFP-adsorberede B. megaterium QM B1551-sporer. Alikvotes af 5 × 108 QM B1551-sporer blev inkuberet med variable koncentrationer af mRFP og blev efterfølgende analyseret ved fasekontrast-, fluorescens- og immunofluorescensmikroskopi som beskrevet i Materialer og metoder. Det samme mikroskopifelt for hver reaktion er rapporteret sammen med sammenlægningspanelet. Eksponeringstiden var 200 ms for alle billeder. Skala bar, 1 μm.
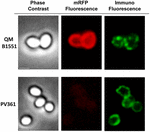
FIGUR 7. Immunofluorescens af mRFP adsorberet til B. megaterium QM B1551 og PV361-sporer. 5 × 108 QM B1551- og PV361-sporer blev inkuberet med 5 μg mRFP og derefter analyseret ved immunofluorescensmikroskopi, som beskrevet i Materialer og metoder. For hvert felt er fasekontrast og immunofluorescensmikroskopi vist. Eksponeringstiden var 200 ms for alle billeder. Skala bar, 1 μm.
Mens QM B1551-sporer, der blev anvendt i eksperimenterne i figur 7, viste en fuldstændig rød fluorescerende ring som i figur 2, viste PV361-sporer et meget svagt rødt fluorescerende signal. Med PV361-sporer blev der kun observeret et rødt signal ved hjælp af lange eksponeringstider i fluorescensmikroskopet (figur 4). Da mRFP er til stede omkring PV361-sporer (figur 3 og 7), konkluderer vi, at mRFP-fluorescensen svækkes, når proteinet er adsorberet til PV361-sporer. Der vil være behov for yderligere eksperimenter for fuldt ud at belyse dette punkt.
Diskussion
De vigtigste resultater af denne rapport er, at sporer af B. megaterium er ekstremt effektive til at adsorbere det heterologe modelprotein mRFP, at exosporiet har en vigtig rolle i denne proces, og at mRFP-molekyler infiltrerer gennem exosporiet og lokaliserer sig mellem den ydre kappe og exosporiet. Disse resultater udvider tidligere arbejde udført på sporer af B. subtilis og viser, at sporer af en anden art også kan bruges til at levere heterologe proteiner ved hjælp af adsorptionsmetoden. Den høje adsorptionseffektivitet, der er observeret med B. megaterium-sporer, skyldes til dels den store størrelse af dens spore sammenlignet med B. subtilis’. B. megaterium-sporens overfladeareal er ca. 2,5 gange større end B. subtilis-sporens med en overflade på 5,33 μm2 (h: 1,60 ± 0,16 w: 0,84 ± 0,07) mod 1,98 μm2 (h: 1,07 ± 0,09 w: 0,48 ± 0,03). De store dimensioner gør det muligt at adsorbere op til 100 μg mRFP, når 160 μg protein reageres med sporer.
Observationen af, at mRFP krydser exosporiet, indikerer, at det er permeabelt for mRFP, et 27 kDa protein. Permeabilitet af exosporiet er ikke helt overraskende, da spirer, der findes i miljøet, skal krydse de ydre lag af sporerne for at nå deres receptorer, om end spirer typisk er små molekyler med molekylmasse typisk <200 Da. Desuden er mRFP-dataene stort set i overensstemmelse med resultaterne af tidligere undersøgelser, der er udført med mærkede dextrans og beslægtede molekyler (Koshikawa et al., 1984; Nishihara et al., 1989). I disse undersøgelser blev B. megaterium QM B1551 exosporium foreslået at udgøre en permeabilitetsbarriere for molekyler med molekylvægte på over 100 KDa, mens det påvirkede passagen af molekyler med en masse et sted mellem 2 og 40 kDa (Koshikawa et al, 1984; Nishihara et al., 1989).
En interessant udfordring for fremtidigt arbejde vil være at fastslå den mekanisme eller infiltrationsvej, som mRFP, og dermed andre heterologe proteiner af interesse, tager for at komme ind i intercoat-rummet. Undersøgelse ved transmissionselektronmikroskopi af afskårne B. megaterium QM B1551-sporer viser, at exosporiet består af to identiske “skaller” (Manetsberger et al., 2015a), og det kan være, at grænsefladen mellem hver af disse strukturer (beskrevet som “apikale åbninger” i tidlige artikler) tillader indtrængen af relativt store molekyler. Det vil også være af interesse at afdække grundlaget for det tilsyneladende tab af mRFP-fluorescens ved adsorption til PV361-sporer, og om mRFP-molekyler er i stand til at infiltrere de ydre kappelag, som observeret for B. subtilis-sporer (Donadio et al., 2016).
I den aktuelle undersøgelse opstiller vi den hypotese, at mRFP-molekyler fortrinsvis krydser exosporium og akkumuleres i intercoat-rummet mellem den ydre kappe og exosporiumet. I denne model bliver mRFP-molekyler først adsorberet og vist på sporeoverfladen, når adsorptionsstederne (eller den volumetriske kapacitet?) i intercoatrummet er tilstrækkeligt optaget. Dette indebærer, at adsorptionsmetoden til overfladevisning kan anvendes med B. megaterium QM B1551-sporer, selv om systemet er afhængigt af forholdet mellem spore og protein, der anvendes i adsorptionsreaktionerne. Da forskellige stammer af B. megaterium længe har været anvendt industrielt til produktion af enzymer såsom amylaser og dehydrogenaser, vitaminer og antimikrobielle molekyler (Vary et al, 2007), tyder vores data på en ny bioteknologisk anvendelse af B. megaterium-sporen som et middel til at binde og levere heterologe proteiner.
Author Contributions
ML og GD – udførte de fleste af eksperimenterne; RA og AS – bidrog til fortolkning af data til arbejdet; GiC, LB og GrC – bidrog til udarbejdelsen af arbejdet og reviderede det kritisk; ER – bidrog til diskussioner og forslag under arbejdet og bidrog til at skrive manuskriptet; RI – ledede arbejdet og bidrog til at skrive manuskriptet. Alle forfattere har læst og godkendt det endelige manuskript.
Funding
Dette arbejde blev støttet af et tilskud (STAR Linea1- 2014) fra Federico II University of Napoli og Compagnia di San Paolo, Italien til RI.
Interessekonflikterklæring
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller finansielle relationer, der kunne opfattes som en potentiel interessekonflikt.
Supplementært materiale
Det supplerende materiale til denne artikel kan findes online på: https://www.frontiersin.org/article/10.3389/fmicb.2016.01752/full#supplementary-material
FIGUR S1 | Hele feltbilleder af fluorescensmikroskopianalysen rapporteret i figur 4. Det samme mikroskopifelt blev observeret ved fasekontrast- og fluorescensmikroskopi. Skala bar 5 μm. Sammenlægningspanelet er rapporteret. Eksponeringstiden er angivet.
FIGUR S2 | Hele feltbilleder af immunofluorescensanalyse, der er rapporteret i figur 7. Det samme mikroskopifelt blev observeret ved fasekontrast- og fluorescensmikroskopi. Skala bar 5 μm. Eksponeringstiden var 200 ms for alle billeder.
TABLE S1 | Densitometrisk analyse af dot blot-eksperimenter med supernatanterne fra adsorptionsreaktionen med QM B1551-sporer (figur 1B).
TABLE S2 | Densitometrisk analyse af dot blot-eksperimenter med supernatanterne fra forskellige vaske efter adsorptionsreaktionen (figur 1C).
TABLE S3 | Densitometrisk analyse af dot blot-eksperimenter med supernatanterne fra adsorptionsreaktionen udført med sporer af stammerne PY79, QM B1551 og PV361 (figur 3B).
TABLE S4 | Densitometrisk analyse af dot blot-eksperimenter med supernatanterne fra adsorptionsreaktionen udført med forskellige mængder mRFP.
Baccigalupi, L., Ricca, E., og Ghelardi, E. (2015). “Non-LAB probiotika: sporeformere,” i Probiotika og præbiotika: Current Research and Future Trends, eds K. Venema and A. P. do Carmo (Norfolk, VA: Caister Academic Press), 93-103. doi: 10.21775/9781910190098.06
CrossRef Full Text | Google Scholar
Campbell, R. E., Tour, O., Palmer, A. E., Steinbach, P. A., Baird, G. S., Zacharias, D. A., et al. (2002). Et monomerisk rødt fluorescerende protein. Proc. Natl. Acad. Sci. U.S.A. 99, 7877-7882. doi: 10.1073/pnas.082243699
PubMed Abstract | CrossRef Full Text | Google Scholar
Cangiano, G., Sirec, T., Panarella, C., Isticato, R., Baccigalupi, L., De Felice, M., et al. (2014). Sps-genprodukterne påvirker spiring, hydrofobicitet og proteinadsorption af Bacillus subtilis-sporer. Appl. Environ. Microbiol. 80, 7293-7302. doi: 10.1128/AEM.02893-14
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutting, S. M. (2011). Bacillus probiotika. Food Microbiol. 28, 214-220. doi: 10.1016/j.fm.2010.03.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Cutting, S., and Vander Horn, P. B. (1990). “Genetic analysis,” in Molecular Biological Methods for Bacillus, eds C. Harwood and S. Cutting (Chichester: John Wiley and Sons), 27-74.
Google Scholar
Díaz-González, F., Milano, M., Olguin-Araneda, V., Pizarro-Cerda, J., Castro-Córdova, P., Tzeng, S. C., et al. (2015). Proteinsammensætning af det yderste exosporielignende lag af Clostridium difficile 630 sporer. J. Proteom. 123, 1-13. doi: 10.1016/j.jprot.2015.03.035
PubMed Abstract | CrossRef Full Text | Google Scholar
Di Luccia, B., D’Apuzzo, E., Varriale, F., Baccigalupi, L., Ricca, E., og Pollice, A. (2016). Bacillus megaterium SF185 inducerer stressveje og påvirker cellecyklusfordelingen i humane tarmepitelceller. Benef. Microb. 7, 609-620. doi: 10.3920/BM2016.0020
PubMed Abstract | CrossRef Full Text | Google Scholar
Donadio, G., Lanzilli, M., Sirec, T., Ricca, E., og Isticato, R. (2016) Protein adsorption og lokalisering på wild type og mutant sporer af Bacillus subtilis. Microb. Cell Fact. 15:153. doi: 10.1186/s12934-016-0551-2
PubMed Abstract | CrossRef Full Text
Eppinger, M., Bunk, B., Johns, M. A., Edirisinghe, J. N., Kutumbaka, K. K. K., Koenig, S. S., et al. (2011). Genomsekvenser af de bioteknologisk vigtige Bacillus megaterium-stammer QM B1551 og DSM319. J. Bacteriol. 193, 4199-4213. doi: 10.1128/JB.00449-11
PubMed Abstract | CrossRef Full Text | Google Scholar
Giglio, R., Fani, R., Isticato, R., De Felice, M., Ricca, E., og Baccigalupi, L. (2011). Organisering og evolution af cotG- og cotH-generne i Bacillus subtilis. J. Bacteriol. 193, 6664-6673. doi: 10.1128/jb.06121-11
PubMed Abstract | CrossRef Full Text | Google Scholar
Harwood, C. R., og Cutting, S. M. (1990). Molekylærbiologiske metoder til Bacillus. Chichester: John Wiley and Sons.
Google Scholar
Huang, J. M., Hong, H. A., Van Tong, H., Hoang, T. H., Brisson, A., og Cutting, S. M. (2010). Mucosal levering af antigener ved hjælp af adsorption til bakteriesporer. Vaccine 28, 1021-1030. doi: 10.1016/j.vaccine.2009.10.127
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., Pelosi, A., De Felice, M., og Ricca, E. (2010). CotE binder sig til CotC og CotU og formidler deres interaktion under dannelse af sporehinde i Bacillus subtilis. J. Bacteriol. 192, 949-954. doi: 10.1128/JB.01408-09
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., Pelosi, A., Zilhão, R., Baccigalupi, L., Henriques, A. O., De Felice, M., et al. (2008). CotC-CotU heterodimerisering under samling af Bacillus subtilis sporehinde. J. Bacteriol. 190, 1267-1275. doi: 10.1128/JB.01425-07
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., og Ricca, E. (2014). Spore overfladevisning. Microbiol. Spectr. 2, 351-366. doi: 10.1128/microbiolspec.TBS-0011-2012
PubMed Abstract | CrossRef Full Text | Google Scholar
Isticato, R., Sirec, T., Treppiccione, L., Maurano, F., De Felice, M., Rossi, M., et al. (2013). Ikke-rekombinant visning af B-underenheden af det varmelabile toksin fra Escherichia coli på vildtype- og mutantsporer af Bacillus subtilis. Microb. Cell Fact. 12:98. doi: 10.1186/1475-2859-12-98
PubMed Abstract | CrossRef Full Text | Google Scholar
Knecht, L. D., Pasini, P., og Daunert, S. (2011). Bakteriesporer som platforme for bioanalytiske og biomedicinske anvendelser. Anal. Bioanal. Chem. 400, 977-989. doi: 10.1007/s00216-011-4835-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Koshikawa, T., Beaman, T. C., Pankratz, H. S., Nakashio, S., Corner, T. R., og Gerhardt, P. (1984). Resistens, spiring og permeabilitetskorrelationer for Bacillus megaterium-sporer, der successivt fjernes fra integumentlagene. J. Bacteriol. 159, 624-632.
PubMed Abstract | Google Scholar
Manetsberger, J., Hall, E. A. H., og Christie, G. (2015a). Plasmidkodede gener påvirker exosporium samling og morfologi i Bacillus megaterium QM B1551 sporer. FEMS Microbiol. Lett. 362:fnv147. doi: 10.1093/femsle/fnv147
PubMed Abstract | CrossRef Full Text | Google Scholar
Manetsberger, J., Manton, J. D., Erdelyi, M. J., Lin, H., Rees, D., Christie, G., et al. (2015b). Ellipsoid lokaliseringsmikroskopi udleder størrelsen og rækkefølgen af proteinlag i Bacillus sporecoats. Biophys. J. 109, 2058-2066. doi: 10.1016/j.bpj.2015.09.023
PubMed Abstract | CrossRef Full Text | Google Scholar
McCloy, R., Rogers, S., Caldon, E., Lorca, T., Castro, A., og Burgess, A. (2014). Delvis hæmning af Cdk1 i G2-fasen tilsidesætter SAC og afkobler mitotiske hændelser. Cell Cycle 13, 1400-1412. doi: 10.4161/cc.28401
PubMed Abstract | CrossRef Full Text | Google Scholar
McKenney, P. T., Driks, A., og Eichemberger, P. (2012). Bacillus subtilis endospore: samling og funktioner af den flerlagede kappe. Nat. Rev. Microbiol. 11, 33-44. doi: 10.1038/nrmicro2921
PubMed Abstract | CrossRef Full Text | Google Scholar
Nicholson, W. L., og Setlow, P. (1990). “Sporulation, germination and out-growth,” in Molecular biological methods for Bacillus, eds C. Harwood and S. Cutting (Chichester: John Wiley and Sons), 391-450.
Google Scholar
Nishihara, T., Takubo, Y., Kawamata, E., Koshikawa, T., Ogaki, J., og Kondo, M. (1989). Den ydre kappes rolle i Bacillus megaterium-sporers resistens. J. Biochem. 106, 270-273.
PubMed Abstract | Google Scholar
Pesce, G., Rusciano, G., Sirec, T., Isticato, R., Sasso, A., og Ricca, E. (2014). Målinger af overfladeladning og hydrodynamisk koefficient af Bacillus subtilis-sporer ved hjælp af optiske pincetter. Colloids Surf. B Biointerfaces 116C, 568-575. doi: 10.1016/j.colsurfb.2014.01.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Ricca, E., Baccigalupi, L., Cangiano, G., De Felice, M., og Isticato, R. (2014). Mucosal vaccineoverførsel ved hjælp af ikke-recombinante sporer af Bacillus subtilis. Microb. Cell Fact. 13, 115. doi: 10.1186/s12934-014-0115-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosso, M. L., og Vary, P. S. (2005). Fordeling af Bacillus megaterium QM B1551 plasmider blandt andre B. megaterium-stammer og Bacillus-arter. Plasmid 53, 205-217. doi: 10.1016/j.plasmid.2004.10.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Rusciano, G. Zito, G., Isticato, R., Sirec, T., Ricca, E., Bailo, E., og Sasso, A. (2014). Nanoskala kemisk billeddannelse af Bacillus subtilis-sporer ved at kombinere spidsforstærket Raman-spredning og avancerede statistiske værktøjer. ACS Nano 8, 12300-12309. doi: 10.1021/nn504595k
PubMed Abstract | CrossRef Full Text | Google Scholar
Sirec, T., Strazzulli, A., Isticato, R., De Felice, M., Moracci, M., og Ricca, E. (2012). Adsorption af beta-galactosidase fra Alicyclobacillus acidocaldarius på sporer af vild type og mutanter af Bacillus subtilis. Microb. Cell Fact. 11:100. doi: 10.1186/1475-2859-11-100
PubMed Abstract | CrossRef Full Text | Google Scholar
Stewart, G. C. (2015). Exosporiumlaget af bakteriesporer: en forbindelse til miljøet og den inficerede vært. Microbiol. Mol. Biol. Rev. 79, 437-457. doi: 10.1128/MMBR.00050-15
PubMed Abstract | CrossRef Full Text | Google Scholar
Vary, P. S., Biedendieck, R., Fuerch, T., Meinhardt, F., Rohde, M., Deckwer, W. -D., et al. (2007). Bacillus megaterium – fra simpel jordbakterie til industriel proteinproduktionsvært. Appl. Microbiol. Biotechnol. 76, 957-967 doi: 10.1007/s00253-007-1089-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Youngman, P., Perkins, J. B., og Losick, R. (1984). En ny metode til hurtig kloning i Escherichia coli af kromosomalt DNA fra Bacillus subtilis ved siden af Tn917-indsættelsen. Mol. Gen. Genet. 195, 424-433. doi: 10.1007/BF00341443
CrossRef Full Text | Google Scholar