T.H. Huxley escribió que mirar a otros simios es ver «copias borrosas de uno mismo» (1863 pág. 73). La estrecha relación física entre los simios, u hominoides, ya había sido reconocida un siglo antes por Carolus Linnaeus (1759). Los hominoideos vivos están unidos por rasgos relacionados con la ortogriedad habitual y los comportamientos por debajo de la rama: torsos anchos con articulaciones de los hombros muy espaciadas, espaldas rígidas, extremidades anteriores largas, articulaciones de las extremidades móviles, fuerte capacidad de agarre y ausencia de cola (Huxley, 1863; Harrison, 1987 & 1991; Shoshani et al., 1996). Los humanos carecen de varias de estas características, como los brazos relativamente largos, como resultado de la adaptación secundaria al bipedismo. Los atributos craneales también unen a los hominoideos, incluyendo paladares anteriores anchos y cerebros relativamente grandes (Jerison, 1973; Rae, 1997). Los hominoideos también tienen una vida prolongada con un retraso en la maduración (Kelley, 1997), y ocupan nichos ecológicos similares: todas las especies, excepto los humanos, habitan en bosques tropicales y dependen en gran medida de la fruta madura y las hojas para alimentarse (Fleagle, 2013).
Los hilobátidos (gibones y siamangs) son más pequeños que otros simios vivos y se diferencian en otros aspectos, lo que refleja su largo periodo de aislamiento evolutivo de los homínidos (grandes simios y humanos), que comparten un ancestro común más reciente (Figura 1). Los homínidos (excepto los humanos) tienen paladares anteriores más ensanchados que los de los hilobátidos (Rae, 1997), poseen espaldas inferiores aún más cortas y rígidas, y carecen de especializaciones anatómicas para la braquiación ricochetal (Young, 2003).
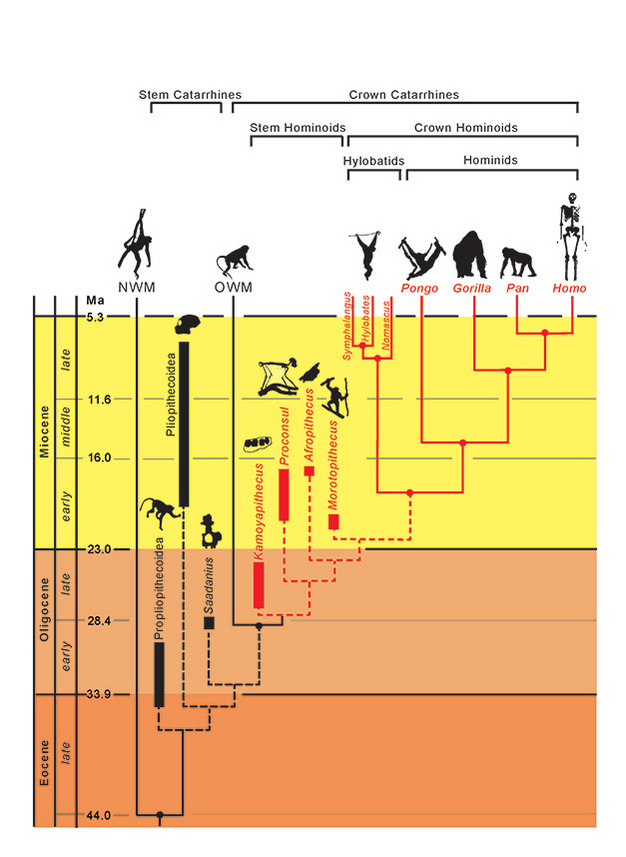
Las relaciones filogenéticas de los taxones están representadas por líneas sólidas (existentes) y discontinuas (extintas). El tiempo geológico se representa en el eje vertical. La cronología de los eventos de ramificación de los taxones es sólo aproximada y se infiere a partir de una combinación de datos moleculares y fósiles (Steiper et al., 2004; MacLatchy et al., 2006; Steiper y Young, 2008 y 2009; Chan et al., 2010; Stevens et al., 2013). Las ubicaciones filogenéticas de los taxones fósiles son hipotéticas, como se discute en el texto. La posición de Afropithecus y Morotopithecus podría invertirse, o ambos podrían ser taxones hermanos. NMW se refiere a los monos del Nuevo Mundo y OWM a los del Viejo Mundo, mientras que Propliopithecoidea, Pliopithecoidea y Saadanius se refieren a los catarrinos de tallo de Afro-Arabia que carecen de cualquier sinapomorfía de los catarrinos de la corona.
Dadas estas similitudes adaptativas entre los hominoideos vivos, es de cierto interés investigar cuál (si es que hay alguna) caracterizó la divergencia inicial de los hominoideos de otros primates catarrinos, especialmente los cercopitecoideos (monos del Viejo Mundo). Las pruebas morfológicas y genéticas combinadas sitúan la divergencia hominoide-cercopitecoide en 29,6 Ma (~24-38 Ma), y la divergencia hilobátido-homínido en 18,8 Ma (~16-24 Ma) (Steiper y Young, 2009). Por tanto, los hominoideos deberían estar presentes en el Oligoceno, pero ¿cómo podemos reconocerlos? ¿Los hemos encontrado? La identificación de estos primeros simios fósiles se ve potencialmente obstaculizada por los siguientes problemas:
1) Un registro fósil deficiente. Hay pocos yacimientos del Oligoceno en África que representen el periodo de tiempo apropiado, y aunque hay varios yacimientos del Mioceno temprano, muchos de sus taxones catarrinos están pobremente representados, particularmente en el postcráneo.
2) El tiempo transcurrido desde el origen de los Hominoidea. Los miembros basales de un taxón pueden tener poco parecido con las formas actuales porque a menudo conservan numerosos rasgos primitivos. Por ejemplo, un simio muy primitivo podría haber sido pronogrado incluso si el último ancestro común de los simios vivos hubiera desarrollado adaptaciones para la postura erguida. Por lo tanto, se espera que las sinapomorfías potenciales entre los hominoideos del tronco y de la corona sean escasas.
3) Los largos periodos de evolución en los linajes existentes pueden haber sido individualmente únicos, de manera que los taxones modernos conservan pocas sinapomorfías definitorias, o algunas de sus presuntas sinapomorfías pueden ser paralelismos.
4) La historia evolutiva de la transformación morfológica puede no estar adecuadamente resuelta, lo que lleva a interpretaciones filogenéticas conflictivas.
El estudio comparativo tanto de los catarrinos del tallo como de la corona puede mitigar estos problemas (Zalmout et al., 2010). Por ejemplo, los rasgos presentes en una especie del Oligoceno tardío o del Mioceno temprano, que no se comparten primitivamente con los catarrinos de tallo y los cercopitecoides, pero que se observan en los hominoideos de la corona, son sinapomorfías plausibles de los hominoideos. Por ejemplo, el seno etmo-frontal se da en los simios africanos y está ausente en los catarrinos de tallo afro-árabes, cercopitecoides, Pongo e Hylobates (Rossie et al., 2002; Rossie, 2008). El seno también aparece en tres hominoideos putativos del Mioceno temprano: Proconsul, Morotopithecus y Afropithecus (ibíd.). La distribución de este rasgo puede interpretarse, por tanto, como un carácter derivado que aparece por primera vez en los hominoideos del tronco y que se pierde en los simios asiáticos (Begun, 1992; Moyà-Solà y Köhler, 1995; pero véase Rossie, 2008, para formas alternativas de interpretar este carácter). Otro rasgo craneal, la mayor robustez transversal de los caninos y la expansión concomitante del paladar anterior, tiene una distribución similar (es decir, se encuentra en los homínidos existentes (perdido secundariamente en los humanos) así como en Proconsul, Morotopithecus y Afropithecus, pero está ausente en los catarrinos de tallo y los cercopitecoides), y también apoya la interpretación de que los taxones del Mioceno son hominoides (Begun et al., 1997; Zalmout et al., 2010). El Kamoyapithecus del Oligoceno tardío sólo se conoce por restos gnatodentales (Leakey et al., 1995), pero la presencia de dientes caninos superiores expandidos transversalmente podría indicar que también es un hominoide (Zalmout et al., 2010). Además, estos cuatro géneros fósiles incluyen especies con tamaños corporales estimados en más de 30 kg, los primeros catarrinos en alcanzar este tamaño.
Dado el gran tamaño y el nicho arbóreo de los simios existentes (los gibones son probablemente enanos en segundo lugar: Tyler, 1993), el gran tamaño corporal puede ser también una característica de los hominoideos de tallo. En los hominoideos de la corona, este rasgo está tradicionalmente ligado a la explotación por parte de los hominoideos de alimentos arbóreos de alta calidad, posibilitada por innovaciones locomotoras que incluyen una mayor movilidad de las extremidades y capacidad de agarre (que permiten distribuir el peso entre múltiples soportes), y la ortogadez (Avis, 1962; Napier, 1967; Tuttle, 1975; Fleagle, 1976; Cachel, 1979; Temerin y Cant, 1983; Cant, 1987; Wheatley, 1987; Kelley, 1997; MacLatchy, 2004).
La evidencia de estas transformaciones postcraneales se expresa de forma variable en Proconsul, Afropithecus y Morotopithecus. Proconsul es bien conocido a partir de varios esqueletos parciales y ha sido reconstruido como un cuadrúpedo de rama superior, con un tronco pronogrado, pero parece haber carecido de cola (Napier y Davis, 1959; Beard et al., 1986; Ward et al., 1991; Ward, 1998). También se ha reconstruido que Proconsul tenía una mayor capacidad de agarre y una mayor movilidad en articulaciones como el codo, en relación con los catarrinos y cercopitecoides primitivos, como demuestran características como una zona conoidea distinta en el húmero distal (Rose, 1988). Afropithecus está menos representado postcranealmente, pero se asemeja a Proconsul en los elementos conocidos (Ward, 1998). Aunque la asociación entre las muestras craneales y postcraneales de las localidades de Moroto es objeto de desacuerdo (Senut et al., 2000), los restos postcraneales de Morotopithecus difieren sistemáticamente de los atribuidos a Proconsul, sugiriendo en su lugar un hominoide con la parte inferior de la espalda rígida, una articulación gleno-humeral móvil y un fémur adaptado para la escalada deliberada, que recuerda más a los hominoides de la corona (MacLatchy et al., 2000).
Además de los tamaños corporales más grandes, todos los taxones que pueden ser evaluados tienen un dimorfismo sexual pronunciado tanto en el tamaño corporal como en la morfología canina (Kelley, 1986; Ruff et al., 1989). Mientras que el fuerte dimorfismo sexual es una característica primitiva de los catarrinos, está más elaborado en los aparentes hominoideos del tronco y en los grandes simios modernos (pero se reduce secundariamente en los humanos).
Hay pocas sinapomorfías, si es que hay alguna, convincentes en la dentición de los hominoideos. Dado que los hominoideos conservan la morfología dental primitiva de los catarrinos (Kay y Ungar, 1997), mientras que los dientes de los cercopitecoideos comparten la clara sinapomorfía de la bilofodoncia molar, existe una historia de uso de rasgos dentales débilmente diagnósticos para reconocer que algunos catarrinos del Oligoceno y del Mioceno poco representados podrían ser hominoideos, a la espera del descubrimiento de especímenes más completos (por ejemplo, Andrews, 1978 y 1992; Stevens et al., 2013). Dichos catarrinos del Oligoceno y del Mioceno han sido denominados informalmente como «simios dentales» (por ejemplo, Fleagle y Kay, 1987; Cartmill y Smith, 2009), o colocados en el limbo taxonómico utilizando una designación como Superfamilia incertae sedis (por ejemplo, Begun, 2007).
En particular, se ha propuesto la idea de que ninguno de los taxones fósiles discutidos anteriormente es un hominoide, y que el primer hominoide reconocible es el Kenyapithecus, de 14 millones de años de antigüedad, que tiene algunos atributos craneodentales derivados (por ejemplo, cíngulo molar reducido, una posición más alta del origen del arco cigomático) en relación con las formas del Mioceno temprano (Harrison, 2010). De ser así, habría una brecha inexplicable de al menos 10 Ma -a pesar de un robusto registro fósil a lo largo de gran parte de este período de tiempo- entre el origen estimado de los hominoideos y su primera aparición.
Aunque los paleoantropólogos no pueden determinar la ubicación filogenética de los «simios dentales» o Kamoyapithecus con confianza, existe un amplio apoyo a la hipótesis de que Proconsul, Afropithecus y Morotopithecus son hominoideos de tallo, en lugar de catarrinos de tallo. Como ventana a la evolución temprana de los hominoideos, estos taxones sugieren lo siguiente:
1) El gran tamaño del cuerpo, la ampliación de la dentición anterior y los sutiles cambios en la dentición posterior (por ejemplo, molares más alargados y cambios en la colocación de las cúspides de los molares y el desarrollo del cíngulo) sugieren la ocupación de nichos ecológicos fundamentalmente diferentes a los de los catarrinos troncales. Los caninos grandes funcionan tanto en la obtención de alimentos como en la selección sexual, con un dimorfismo pronunciado que sugiere que esto último fue un factor. El gran tamaño permite el acceso a nuevos alimentos y mejora la flexibilidad del forrajeo (Wheatley, 1987), disminuye los riesgos de depredación, favorece la supervivencia y, unido a factores como la estabilidad del hábitat, puede estar relacionado con el retraso de la maduración. El análisis del patrón de desarrollo dental de Afropithecus (y posiblemente de Proconsul) sugiere un retraso en la maduración con respecto a otros antropoides (Kelley, 1997, 2002), lo que hace que la historia de vida lenta sea otra posible sinapomorfía de los hominoideos.
2) A principios del Mioceno, existe al menos una modesta diversidad locomotora y dietética entre los probables hominoideos de cuerpo grande, lo que indica el comienzo de una verdadera radiación adaptativa, con un tamaño corporal cada vez mayor, una mayor novedad locomotora y una explotación más especializada de las fuentes de alimento arbóreas que evolucionan simultáneamente.
3) Actualmente no se puede resolver qué rasgos anatómicos asociados con la ortogriedad y la movilidad articular en los simios modernos son homólogos u homoplásticos (por ejemplo, Harrison, 1991; Larson, 1998; Young, 2003; MacLatchy, 2004 & 2010; Lovejoy et al., 2009). Este debate tiene implicaciones para interpretar la posición evolutiva de los supuestos hominoides troncales, y para reconstruir el patrón y el momento de la aparición de las adaptaciones de los simios modernos.
Por ejemplo, las similitudes postcraneales entre Morotopithecus y los simios vivos podrían interpretarse de varias maneras. Si los simios vivos adquirieron su ortogonalidad a través de una evolución paralela, entonces el Morotopithecus representa el primer caso conocido de una transición que ocurrió varias veces en la evolución de los hominoideos. Esto es de gran interés adaptativo, ya que sugiere que alguna propensión genética o de desarrollo subyacente (véase más adelante) siguió dirigiendo la respuesta evolutiva de los hominoideos en la misma dirección. También significaría que ciertos rasgos postcraneales deben utilizarse con precaución en los análisis filogenéticos (Sanders y Bodenbender, 1994). Por otro lado, si estas similitudes locomotoras fueran heredadas de un ancestro común, Morotopithecus estaría más estrechamente relacionado con los hominoideos coronados que Proconsul (y posiblemente Afropithecus). Otra posibilidad es que Morotopithecus fuera un «experimento» independiente, y que el ancestro común de los hominoideos coronados fuera, sin embargo, ortógrado.
4) Como se ha discutido anteriormente, a pesar de las diferencias en las especializaciones locomotoras y de los largos periodos de evolución independiente, todos los taxones hominoideos supervivientes comparten un plan corporal similar que los diferencia de otros primates (con la excepción de algunos monos atelinos, que han evolucionado convergentemente torsos y especializaciones de las extremidades anteriores similares a las de los simios (Erikson, 1963; Larson, 1998). Los fundamentos anatómicos básicos del plan corporal de los simios han dado lugar a una notable diversidad y versatilidad locomotora, lo que ha permitido a los primates de gran tamaño desplazarse eficazmente de forma arbórea (mediante la escalada y la suspensión) y terrestre (utilizando la marcha con nudillos y el bipedismo). Esta innovación locomotora, ligada a características como la ortogonalidad, probablemente sea la base del «éxito» evolutivo de todos los hominoides vivos, y parece haber estado presente en cierta medida en los hominoides troncales. El trabajo de desarrollo de Young y sus colegas (2010) ha demostrado que los simios actuales tienen una integración reducida entre las extremidades anteriores y posteriores en comparación con los monos cuadrúpedos. Se ha interpretado que la relajación de esta restricción genética ha permitido que las extremidades evolucionen más libremente de diferentes maneras en respuesta a las presiones de selección (ibíd.). El uso diferencial inferido de las extremidades en Morotopithecus (MacLatchy et al., 2013) y en taxones más jóvenes del Mioceno (por ejemplo, Moya Sola y Kohler, 1996; Moya Sola et al, 2004; Nakatsukasa y Kunimatsu, 2009) apoya provisionalmente la suposición de Young y sus colegas (2010) de que la «evolucionabilidad» de las extremidades puede ser un rasgo ancestral de los hominoideos, pero se necesita más material fósil y una mejor comprensión de la genética que subyace a este rasgo.
La comprensión futura de los orígenes de los hominoideos y de su evolución temprana surgirá de los continuos intentos de identificar y comprender la base de las posibles sinapomorfías de los hominoideos. Esto, a su vez, requerirá un mejor registro fósil tanto de los catarrinos del tronco como de la corona, y un estudio anatómico, de desarrollo y genético más detallado de los caracteres compartidos que se están analizando. El estudio integrado de los hominoides fósiles en su contexto paleoambiental también puede ayudarnos a comprender mejor por qué se seleccionaron los rasgos hominoides, ya sea una o varias veces.